| Суббота, 09.05.2026, 04:56 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В категории материалов: 23
Показано материалов: 1-10 |
Страницы: 1 2 3 » |
Сортировать по:
Дате ·
Названию ·
Комментариям ·
Просмотрам
Рассматривая
круговорот углерода, мы уже противопоставляли друг другу два процесса: фотосинтез, при котором фиксируется СО2 и
выделяется О 2, и минерализацию органических
веществ, связанную с потреблением О2 и освобождением СО2.
Процессы эти противоположны. С точки зрения массообмена важнее всего переход
углерода из газообразного неорганического вещества в полутвердые и твердые
органические соединения и обращение этого процесса. Если же рассматривать оба
этих процесса с позиций энергообмена, т.е. преобразования энергии, то углерод
имеет меньшее значение, чем водород. |
Метаболизм
и метаболические пути. Как во время роста, так и в состоянии покоя
вегетативные клетки нуждаются в постоянном притоке энергии. Живая клетка
представляет собой высокоорганизованную материю. Энергия необходима не только
для создания такой организации, но и для ее поддержания. Эту энергию организм
получает в процессе обмена веществ, или метаболизма, т.е. путем регулируемых
превращений, которым различные вещества подвергаются внутри клеток. Источниками
энергии служат питательные вещества, поступающие из внешней среды. В клетках
эти вещества претерпевают ряд изменений в результате последовательных
ферментативных реакций, образующих этапы определенных метаболических путей.
Такие пути выполняют две главные функции: они, во-первых, поставляют материалы
- предшественники для построения клеточных компонентов и, во-вторых,
обеспечивают энергию для клеточных синтезов и других процессов,
требующих затраты энергии. |
Несколько
путей ведут от глюкозы к С3-соединениям и среди них к пирувату - одному
из важнейших промежуточных продуктов метаболизма. Чаще других используется путь
распада через образование фруктозе-1,6-дисфосфата; его называют фруктозодисфосфатным
путем, гликолитическим расщеплением, гликолизом или (по имени изучивших его
исследователей) путем Эмбдена-Мейергофа-Парнаса (рис. 7.3). 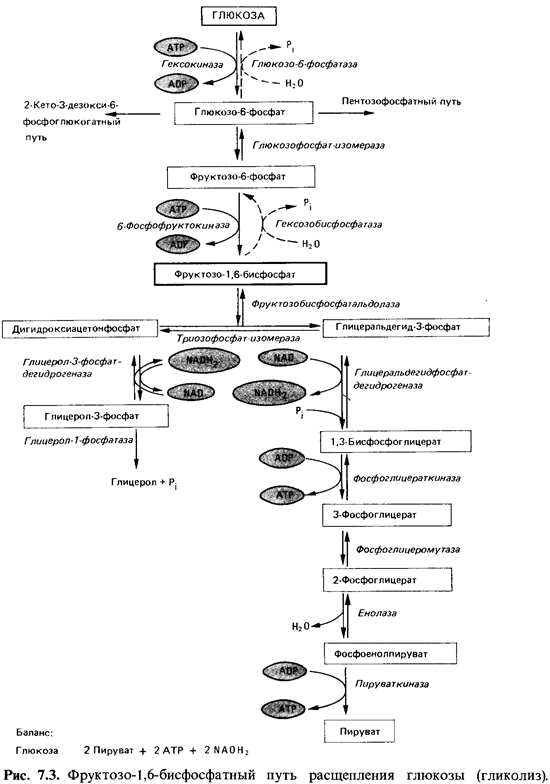
Другой ряд реакций,
к осуществлению которых способно большинство организмов, образует цикл,
известный под названием окислительного пентозофосфатного пути,
гексозомонофосфатного пути или схемы Варбурга-Диккенда-Хореккера (рис. 7.4).
Обратная последовательность; реакций этого пути включает важные
этапы, ведущие к регенерации акцептора СО2 при автотрофной фиксации
углекислоты. Только у бактерий встречается, видимо, путь Энтнера-Дудорова, или,
как его еще называют, КДФГ-путь (по характерному промежуточному
продукту-2-кето-3-дезокси-6-фосфоглюконату, КДФГ; рис. 7.5). Другие сходные
механизмы распада гексоз имеют более специальное значение. |
На фруктозодисфосфатном
пути (рис. 7.3) глюкозо-6-фосфат при подготовке к расщеплению изомеризуется глюкозофосфат-изомеразой в
фруктозо-6-фосфат; затем происходит фосфорилирование в положении 1 под
действием фосфофруктокиназы за счет АТР. Образовавшийся фруктозе-
1,6-дисфосфат расщепляется фруктозодисфосфатальдолазой до дигидроксиацетонфосфата
и глицеральдегид-3-фосфата. Оба триозофосфата находятся в равновесии между
собой; установление этого равновесия катализируется триозофосфат-изомеразой. Дигидроксиацетонфосфат
может восстанавливаться глицеролфосфат-дегидрогеназой до глицеролфосфата,
который гидролизуется глицерол-1-фосфатазой с образованием глицерола и
ортофосфата. Обычно же сначала происходит превращение образовавшегося под
действием альдолазы дигидроксиацетонфосфата в глицеральдегид-3-фосфат, который
затем окисляется. |
В
пентозофосфатном пути (рис. 7.4) глюкозо-6-фосфат дегидрируется глюкозо-6-фосфат-дегидрогеназой;
при этом водород переносится на NADP и образуется 6-фосфоглюконолактон, который
спонтанно или при участии фермента (глюконолактоназы) гидролизуется до
6-фосфоглюконата. Этот последний дегидрируется дегидрогеназой до З-кето-6-фосфоглюконата,
из которого затем путем декарбоксилирования образуется рибулозо-5-фосфат. Этим
завершается собственно процесс окисления. |
Глюкозо-6-фосфат
сначала, как это было описано выше для пентозофосфатного пути, дегидрируется до
6-фосфоглюконата. Под действием фосфоглюконатдегидратазы от него
отщепляется вода и образуется 2-кето-3-дезокси-6-фосфоглюконат (рис.
7.5). Кетодезоксифосфоглюконат расщепляется специфической альдолазой на пируват
и глицеральдегид-3-фосфат. Последний окисляется до пирувата, так же как и в
фруктозодисфосфатном пути. |
Пируват
занимает центральное положение в промежуточном метаболизме и может служить
предшественником разнообразных продуктов. Многие организмы окисляют большую
часть образующегося в ходе катаболизма пирувата до ацетилкофермента А.
Важнейшую роль у бактерий играют три реакции:
- (1) Пируват + СоА + NAD -> Ацетил-СоА + NADH2 +
СО2
- (2) Пируват + СоА + 2Fd -> Ацетил-СоА + 2FdH + СО2
- (3) Пируват + СоА -> Ацетил-СоА + Формиат
|
Цикл
трикарбоновых кислот (рис. 7.7) служит для окисления двухуглеродного соединения
ацетата до СО2 с отщеплением водорода. При участии трех
дегидрогеназ водород переносится на NAD(P), а под действием сукцинатдегидрогеназы
- непосредственно на хинон. Как правило, коферменты передают водород в
дыхательную цепь. |
В то время
как большинство анаэробных организмов способно синтезировать АТР только путем
фосфорилирования на уровне субстрата, аэробы могут осуществлять несравненно
более эффективную регенерацию АТР. Они обладают особым аппаратом: дыхательной
(электрон-транс портной) цепью и ферментом ATР-синтазой;
обе системы у прокариот находятся в плазматической мембране, а у эукариот – во
внутренней мембране митохондрий. Ведущие свое происхождение от субстратов
восстановительные эквиваленты (Н или электроны) в этих мембранах по ступают в
дыхательную цепь, и электроны переносятся на О2 (или другие
терминальные акцепторы электронов). В дыхательной цепи происходят реакции,
представляющие собой биохимический аналог сгорания водорода. От химического
горения молекулярного водорода они отличаются тем, что значительная часть
свободной энергии переводится при этом в биологически доступную форму, т. е. в
АТР, и лишь небольшая доля рассеивается в виде тепла. |
Транспорт
водорода и транспорт электронов - процессы эквивалентные. Дыхательная цепь
может рассматриваться как цепь переноса электронов. Компоненты дыхательной цепи
переходят попеременно из окисленного состояния в восстановленное и обратно, т.
е. ведут себя как типичные окислительно-восстановительные катализаторы. Они
обладают окислительно-восстановительным потенциалом, который может быть измерен
непосредствен но (у цитохромов) или косвенно (у NAD, FAD). |
|
 |  |


