| Суббота, 09.05.2026, 04:56 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В категории материалов: 10
Показано материалов: 1-10 |
|
Сортировать по:
Дате ·
Названию ·
Комментариям ·
Просмотрам
Способность
использовать свет как источник энергии, необходимой для роста,
присуща двум группам бактерий, принципиально отличающимся друг от друга.
Пурпурные
и зеленые бактерии, объединяемые в порядок Rho-dospirillales1,
можно рассматривать как реликтовые организмы, дошед шие до нас из времен
начальной эволюции фотосинтеза. Они не в со стоянии использовать в качестве
донора водорода воду (как это делают зеленые растения); им требуются доноры с
более высокой степенью восстановления (H2S, H2 или
органические вещества). Поэтому фотосин тез у этих бактерий протекает без
выделения 02. В таких случаях гово рят об аноксигенном фотосинтезе.
Бактерии этой группы - типичные во дные организмы, распространенные как в
пресной, так и в морской воде. Их красная, оранжевая или зеленая окраска
обусловлена присут ствием бактериохлорофиллов и каротиноидов. |
Фототрофные
бактерии, осуществляющие аноксигенный фотосинтез, де лятся на две большие
группы: пурпурные бактерии (Rhodospirillales) и зеленые бактерии
(Chlorobiales). Представители этих двух порядков значительно различаются по
своим цитологическим и физиологическим признакам, а также по характерным для
них пигментам. |
Благодаря
фотосинтетическим пигментам достаточно густые суспензии фототрофных бактерий
имеют зеленую, сине-зеленую, пурпурно-фиоле товую, красную, коричневую или
розовую окраску. Цвет зависит от природы и количественного соотношения
пигментов. Отдельные пиг менты можно распознать даже по спектрам поглощения
интактных клеток (рис. 12.10). Хлорофиллы, например, ответственны за максимумы
поглощения в синей (< 450 нм) и в красной и инфракрасной (650-1100 нм)
областях спектра. Поглощение в области 400-550 нм обусловлено главным образом
каротиноидами, а у цианобактерий в области 550-650 нм-фикобилипротеинами. 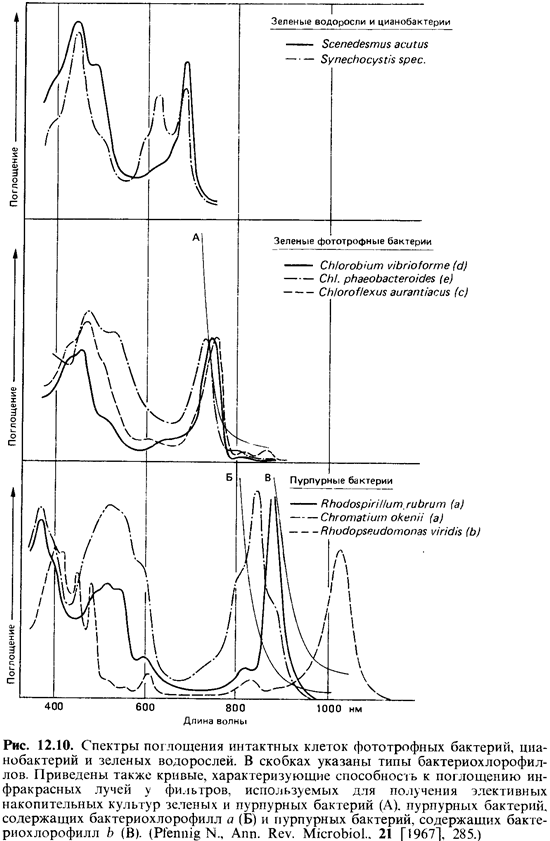
|
Исследование
обмена веществ у фототрофных бактерий сопряжено с ря дом трудностей. Эти
бактерии часто отличаются большой разносторон ностью. Например, многие несерные
пурпурные бактерии способны как к анаэробному росту на свету, так и к аэробному
росту в темноте; дру гие группы представлены строгими анаэробами и облигатными
фото-трофами. Многие виды используют в качестве донора электронов молекулярный
водород, а некоторые-сероводород или серу. Наблюдается ассимиляция С02 и
органических веществ. В анаэробных условиях в темноте за счет брожения может
получаться небольшое количество энергии, недостаточное, однако, для роста
большинства видов. Короче говоря, фототрофные бактерии относятся к организмам,
наиболее раз носторонним в отношении обмена веществ. Поэтому здесь мы можем
лишь глобально охарактеризовать их физиологические
возможности. |
Фототрофные
пурпурные и зеленые бактерии обитают в анаэробных зо нах многих водоемов, в
мелководных прудах, медленно текущих водах, в озерах и морских бухтах.
Пурпурные серобактерии нередко образуют налеты, окрашенные во всевозможные
оттенки красного цвета, от неж но-розового до темно-красного, на поверхности
ила или на каком-ни будь разлагающемся растительном материале. Иногда они
роятся над поверхностью ила, образуя слой толщиной около дециметра. Такое
«цветение» воды в мелких водоемах (рис. 12.12) вызывают в первую очередь
крупные представители пурпурных бактерий - Chromatium okeniu С.warmingii,
С. weissei и Thiospirillum jenense, однако
встречаются и мел кие представители Chromatiaceae и Chlorobiaceae. Бурное
размножение пурпурных серобактерий наблюдается и в таких мелководных прудах,
поверхность которых покрыта плотным слоем ряски (Lemna) или же листьями водяных
лилий. Этот своеобразный биологический фильтр по глощает те спектральные
компоненты света, которые могли бы исполь зоваться зелеными водорослями и
цианобактериями, но пропускает свет, поглощаемый бактериохлорофиллами и
темно-красными кароти-ноидами. Поэтому под покровом из ряски растут анаэробные
фото трофные бактерии, но не развиваются зеленые водоросли и цианобакте-рии
(рис. 12.12). Некоторые виды фототрофных серобактерий предста влены
в естественных местообитаниях почти чистой культурой. Всегда можно обнаружить и
Rhodospirillaceae, но лишь редко в столь массовом количестве. |
Метод отбора
анаэробных фототрофных бактерий основан на том (опи санном выше) наблюдении,
что пурпурные серные бактерии достигают иногда массового развития и в мелких
водоемах, если вся поверхность воды покрыта плотным слоем ряски. С помощью
фильтров, поглощаю щих коротковолновую часть спектра и пропускающих
инфракрасные лучи, используемые лишь отдельными группами фототрофных бакте рий,
можно создать селективные условия для роста как зеленых серо бактерий, так и
пурпурных бактерий, содержащих бактериохлорофилл а или b. |
Под
фотосинтезом понимают происходящее в клетках фототрофных организмов преобразование
световой энергии в биохимически доступную энергию (АТР) и восстановительную
силу [NAD(P)H2], а также свя занный с этим синтез клеточных
компонентов. Фотосинтетическое фос-форилирование и фотосинтетическое
восстановление пиридиннуклеоти-да-это процессы, ведущие к образованию первых стабильных
продук тов фотосинтеза.
К
такому пониманию пришли в результате экспериментальных и теоретиче ских
исследований, основанных главным образом на сравнении фотосинтеза у фототрофных
бактерий и у зеленых растений. После того как Виноградский (1888) установил,
что для некоторых бактерий источником энергии при ассими ляции С02отнюдь
не всегда служит свет, а Энгельман (1883-1888) на основании своих
физиологических исследований отнес пурпурные бактерии к фототрофныморганизмам,
Будер (1919) показал, что эта группа бактерий обладает новым, не известным
ранее типом метаболизма. Серные и несерные пурпурные бактерии ассимилируют С02 или
органические вещества на свету. Однако такого рода фотосинтез существенно
отличается от фотосинтеза высших растений: 1) вода здесь не может служить
донором водорода, и поэтому такой фотосинтез не со провождается выделением 02;
2) бактерии используют в качестве доноров водо рода не воду, a H2S
или органические вещества. |
Первичные
процессы фотосинтеза протекают в тилакоидах - плоских
замкнутых мембранных пузырьках, содержащихся в клетках цианобак терий и в
хлоропластах водорослей и высших растений.
Тилакоидные
мембраны и светособирающие пигменты (пигменты антенн). Тилакоидная
мембрана содержит в себе пигментные молекулы (хлорофилл а, хлорофилл b и каротиноиды),
переносчики электронов и ферменты. Подавляющее большинство молекул хлорофилла (
> 99,5%), а также дополнительные пигменты (каротиноиды, фикобили-протеины)
ответственны за поглощение света и распределение энергии; они образуют систему
антенны. Лишь незначительная часть хлорофил ла авыполняет роль
фотохимического реакционного центра, в котором протекает собственно
фотохимическая окислительно-восстановительная реакция. Пигменты антенн
(светособирающие пигменты) улавливают свет и передают энергию хлорофиллу
реакционного центра (Каротиноид -> Каротиноид*; Хлорофилл + Каротиноид*
-> Хлоро филл* + Каротиноид). Каротиноиды выполняют также защитную функцию:
при очень ярком солнечном освещении они отдают избыточ ную энергию в окружающую
среду и тем самым защищают молекулы хлорофилла от фотоокисления. Система
светособирающих пигментов и реакционный центр объединены в так называемую
фотосинтетиче скую единицу. |
Фотосинтетический
транспорт электронов у анаэробных фототрофных бактерий во многих отношениях
отличается от только что описанного. В аноксигенном фотосинтезе участвует
только одна световая реакция; она поддерживает циклический транспорт
электронов. Электроны, поки дающие цикл для восстановления NAD, не являются
продуктом разло жения воды. Фотосинтез зависит от наличия в среде
восстановленных субстратов и не сопровождается выделением 02.
Собственно фотореак ция хотя и аналогична первой фотореакции у зеленых
растений, однако у некоторых бактерий она приводит, вероятно, лишь к созданию
про тонного потенциала и тем самым к запасанию энергии (АТР), но не к
восстановлению NAD. Таким образом, нециклический перенос элек тронов (от донора
электронов к пиридиннуклеотиду) здесь отсутствует. По-видимому, NADH2 образуется
в результате какой-то темновой реак ции в ходе обратного транспорта электронов,
протекающего с затратой энергии.
Следует,
однако, заметить, что у фототрофных бактерий между от дельными группами
существуют гораздо большие различия в составе пигментов и механизмах
фотосинтеза, чем у зеленых растений. В после дующем изложении мы вначале не
будем касаться зеленых бактерий. |
Виды,
относящиеся к роду Halobacterium (Н. halobium, H. cutirubrum), со
ставляют высокоспециализированную в физиологическом отношении группу бактерий.
Места их естественного обитания-высококонцентрированные или насыщенные
растворы солей, такие как воды Большого соленого озера в США или места, где
добывают соль путем выпарива ния морской воды. Приспособленность бактерий к
существованию в столь экстремальных условиях связана с тем, что концентрация
соли внутри клеток так же высока, как и в окружающей среде. Оптимальный рост
таких бактерий наблюдается в 3,5-5,0
М растворе NaCl. Для нор мального функционирования
выделенных из клеток ферментов тоже не обходимы более чем двухмолярные
концентрации соли. Галобактерии относятся к архебактериям. |
|
 |  |


