| Среда, 13.05.2026, 04:02 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В разделе материалов: 204
Показано материалов: 81-90 |
Страницы: « 1 2 ... 7 8 9 10 11 ... 20 21 » |
Физиологическую
группу бактерий, восстанавливающих сульфат (их называют также
десульфатирующими, сульфатредуцирующими или сульфидобразующими), отличает
способность к переносу водорода с субстрата на сульфат как конечный акцептор
электронов и, таким образом, к восстановлению сульфата до сульфида. В этом
процессе происходит перенос электронов, и в нем участвует цитохром с. Энергия
запасается благодаря фосфорилированию в электрон-транспортной цепи в анаэробных условиях:
Поскольку
такое восстановление сульфата обладает формальным сходством с дыханием, при
котором акцептором водорода служит кислород, принято говорить о сульфатном
дыхании, или о диссимиляционной сульфатредукции. Главным
продуктом такого процесса является сероводород:
8[Н]
+ SO*" -» H2S
+ 2Н20 + 20Н-
Большая часть
сероводорода, образующегося в природе, возникает благодаря этой реакции.
Сульфатредуцирующие бактерии являются, в отличие от нитратредуцирующих,
облигатными анаэробами, т.е. нуждаются в строго анаэробных условиях. |
Давно
известно, что суспензия дрожжей, сбраживающая сахар, вовлекает добавленный к
ней серный цвет в «вихрь биологических восстано влений» с образованием
сероводорода. Однако лишь недавно выясни лось, что и некоторые бактерии
способны расти в присутствии элементарной серы, используя ее в качестве
акцептора водорода при анаэробном переносе электронов. Сера, восстанавливается
при этом до сероводорода. Этот процесс можно назвать серным дыханием. |
Метан
образуется при анаэробном разложении органических веществ. Его запасы весьма
значительны; примерные подсчеты показывают, что 1-1,5% углерода, поступающего в
результате минерализации органиче ских веществ в виде С02 в
атмосферу, сначала попадает туда в виде ме тана и лишь затем превращается под
воздействием гидроксильных ра дикалов (ОН') в СО, а потом в С02. К
экосистемам, в которых образуется метан, относятся большие площади, занятые
тундрой и бо лотами (отсюда другое название метана-болотный газ); рисовые поля;
осадки на дне прудов и озер; лиманы, марши и эстуарии; отстойники очистных
сооружений и, наконец, желудки (рубцы) более чем 109 жвачных
животных. В анаэробных условиях органические вещества сна чала через ряд
промежуточных этапов сбраживаются до ацетата, С02 и Н2.
Эти продукты метаболизма первичных и вторичных деструкторов используются
метанобразующими (метаногенными) бактериями. |
В различных
местах, где образуется метан, происходит также образова ние уксусной кислоты.
Подкисление среды в отстойниках водоочистных сооружений, вероятно, связано с
деятельностью бактерий, превращаю-молекулярный водород
в уксусную кислоту согласно уравнению
4Н2
+ 2С02 -> СН3-СООН
+ 2Н20; AG0 =-111 кДж/моль (- 26,6
ккал/моль) |
Мы уже
неоднократно говорили о сукцинате (янтарной кислоте) как о продукте брожения,
но не упоминали о том, что образование сукцина та может быть связано с
окислительным фосфорилированием. Сукцинат является продуктом восстановления
фумарата:
2[Н]
+ Фумарат -> Сукцинат
Функция
фумарата не сводится к роли простого акцептора, взаимо действующего с NADH2,
который образуется при окислении гексоз. Для пары фумарат/сукцинат
окислительно-восстановительный потен циалEq = - 30 мВ. Фумарат
может акцептировать электроны, которые поставляются переносящими водород
коферментами и уже прошли часть пути по дыхательной цепи; поэтому он делает
возможным окис лительное фосфорилирование. Такого рода фосфорилирование с
фума-ратом в качестве конечного
акцептора электронов можно отнести к
«анаэробным дыхательным процессам» и говорить о фумаратном дыхании (рис.
9.8). 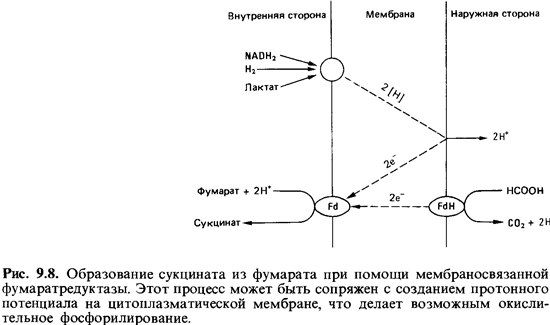
|
Смешанные
популяции почвенных бактерий в анаэробных условиях вос станавливают ионы Fe(III) до
Fe(II). Если в среде помимо Fe(III) при сутствуют также ионы
нитрата и нитрита, то сначала восстанавливают ся они (до нитрита и N2,
денитрификация) и лишь после этого-ионы Fe(III). Предполагают,
что перенос электронов на трехвалентное железо осуществляет нитратредуктаза А.
Поскольку восстановление нитрата сопряжено с окислительным фосфорилированием,
не исключено, что и восстановление Fe (III) может
использоваться в процессе «анаэробного дыхания». Окислительно-восстановительный
потенциал £0', который для пары Fe3 +/Fe2 + равен
+ 770 мВ, делает такую реакцию термодинами чески возможной. Поскольку оксиды
трехвалентного железа практиче ски нерастворимы в воде, они сначала должны быть
переведены в растворимую форму, способную проникать внутрь бактериальных
клеток. Это, вероятно, осуществляется с помощью сидерофоров. Неудивитель но,
что в таких условиях наблюдается лишь медленный и незначительный рост бактерий. |
Большинство
аэробных микроорганизмов окисляет органические пита тельные вещества в процессе
дыхания до С02 и воды. Поскольку в мо лекуле СО 2 достигается
высшая степень окисления углерода, в этом случае говорят о полном
окислении и отличают этот тип дыхания от не полных окислений, при
которых в качестве продуктов обмена выделяют ся частично окисленные органические
соединения.
Под «полным
окислением» имеется в виду лишь то, что не происходит вы деления каких-либо
органических веществ; но это вовсе не означает, что окис ляется весь
поглощенный субстрат. В каждом случае значительная часть суб страта (40-70%) ассимилируется,
т.е. превращается в вещества клеток.
Конечными
продуктами «неполных окислений» могут быть уксусная, глюконовая, фумаровая,
лимонная, молочная кислоты и ряд других со единений. Поскольку эти продукты
сходны с теми, которые образуются при брожениях (пропионовая, масляная,
янтарная, молочная кислоты и др.), а также в связи с тем, что при промышленных
процессах броже ния необходимы специальные технические устройства (ферментеры),
неполные окисления называют также «окислительным брожением» или «аэробной
ферментацией». Слова «брожение» и «ферментация» в этом случае отражают скорее
технологический аспект.
Мы будем
причислять к «неполным окислениям» также простое от щепление водорода от
субстрата и использование микроорганизмов для катализа некоторых реакций, не
имеющих для них какого-либо значе ния в обычных условиях. Ниже будут
рассмотрены некоторые примеры таких окислений. |
Всем
уксуснокислым бактериям присуща способность к образованию кислот путем
неполного окисления Сахаров или спиртов. Эти кислоты как промежуточные или как
непригодные для использования конечные продукты обмена выделяются в среду.
Уксуснокислые бактерии-это грам-отрицательные палочки с перитрихально (Acetobacter) или
полярно (Acetomonas = Gluconobacter) расположенными
жгутиками, обладающие некоторой подвижностью. Они сходны с псевдомонадами, но
отличают ся от них высокой толерантностью к кислотам, слабой пептолитической
активностью, малой подвижностью и отсутствием пигментов. В при родных условиях
уксуснокислые бактерии поселяются обычно на расте ниях. В сахаристых выделениях
растений вместе с уксуснокислыми бак териями обитают дрожжи. |
Многие
органические кислоты, в том числе и аминокислоты, получают в промышленных
масштабах с помощью микроорганизмов, осущест вляющих неполные окисления.
Преимущества использования грибов для получения лимонной, итаконовой,
глюконовой, яблочной и других кислот выяснились уже в начале нашего столетия.
Что касается бакте рий, то еще десять лет назад с их помощью получали только
уксусную и глюконовую кислоты; однако теперь они играют большую роль и как
продуценты аминокислот. |
Для грибов
характерен окислительный тип метаболизма. Это не озна чает, что грибы не
способны к анаэробному расщеплению углеводов, т.е. не могут их сбраживать (ведь
спиртовое брожение осуществляется как раз дрожжами!); однако в анаэробных условиях
сколько-нибудь длительный рост грибов невозможен. Кроме того, основными продук
тами брожения оказываются этанол или молочная кислота. Другие ор ганические
кислоты образуются только в аэробных условиях.
В
естественных местах обитания грибов, т. е. в почве, никогда не бы вает
заметного накопления промежуточных продуктов их жизнедеятель ности. При
недостатке питательных веществ грибы получают максимум энергии и образуют
клеточные вещества за счет полного окисления и ассимиляции субстрата. Если в
лабораторных условиях и в промыш ленной практике дело обстоит иначе и
многочисленные продукты обме на выделяются в среду, то это следует объяснить
избыточным снабже нием грибов углеводами и известной «дезорганизацией» их
метаболиз ма, которой часто способствует исключение из среды некоторых
микроэлементов.
Известно, что
грибы обладают мощной гликолитической системой. В «узких местах» метаболических
путей происходит накопление проме жуточных продуктов, которые прямо или после
назначительных хими ческих изменений выделяются в среду. Дж. Фостер особо
подчеркивал отклонения в обмене веществ, возникающие при избытке субстрата. В
конечном счете большинство неполных окислений связано с наруше нием регуляции
метаболизма. |
|
 |  |


