Мы уже
неоднократно говорили о сукцинате (янтарной кислоте) как о продукте брожения,
но не упоминали о том, что образование сукцина та может быть связано с
окислительным фосфорилированием. Сукцинат является продуктом восстановления
фумарата:
2[Н]
+ Фумарат -> Сукцинат
Функция
фумарата не сводится к роли простого акцептора, взаимо действующего с NADH2,
который образуется при окислении гексоз. Для пары фумарат/сукцинат
окислительно-восстановительный потен циалEq = - 30 мВ. Фумарат
может акцептировать электроны, которые поставляются переносящими водород
коферментами и уже прошли часть пути по дыхательной цепи; поэтому он делает
возможным окис лительное фосфорилирование. Такого рода фосфорилирование с
фума-ратом в качестве конечного
акцептора электронов можно отнести к
«анаэробным дыхательным процессам» и говорить о фумаратном дыхании (рис.
9.8). 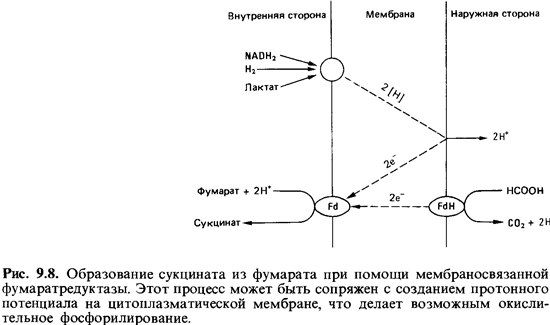
Фумаратное
дыхание широко распространено у хемоорганотрофных анаэробных бактерий.
Добавление фумарата к питательной среде вызы вает у многих бактерий быстрый
рост и повышает выход биомассы. Это указывает на то, что фумарат делает
возможной эффективную реге нерацию АТР. Соответствующие результаты были
получены для энте-робактерий (Escherichia, Proteus, Salmonella, Klebsiella), а
такжеBacteroides, Propionibacterium и Vibrio succinogenes (см.
рис. 8.8). В анаэ робных условиях фумарат, добавленный в питательную среду,
может служить для бактерий экзогенным конечным акцептором электронов. Однако
возможно и его эндогенное образование из углеводов и многих других субстратов
через оксалоацетат и малат (см. рис. 7.13).
Локализация
фумаратредуктазы в клетке согласуется с представлением об ее роли в транспорте
электронов: этот фермент связан с мембраной. Процесс восстановления фумарата
сопровождается затратой протонов. Из гипотезы, согласно которой восстановление
фумарата ведет к созданию про тонного потенциала (с положительным зарядом
снаружи), вытекает предположе ние, что фермент фумаратредуктаза должен
находиться на внутренней поверхно сти мембраны. По-видимому, так и обстоит
дело. Измерения показали, что при восстановлении 1 моля фумарата
образуется 1 моль АТР.
Можно
полагать, что в большинстве процессов брожения, связанных с образованием
сукцината, участвует фумаратредуктаза, и это приводит к дополнительному
образованию АТР.
К таким
процессам относится, например, образование сукцината у Escherichia coli и
пропионата уPropionibacterium. Однако «фумаратное дыха ние»
встречается не только у прокариот. Существует несколько видов
факультативно-анаэробных червей (Ascaris lumbricoides, Fasciola hepatica, Trichurisvulpis, Arenicola marina), способных
жить в анаэробных условиях. Эти организмы выде ляют сукцинат и пропионат;
образование пропионата идет по метилмалонил-СоА-пути (см. рис. 8.3). Роль
образования сукцината и пропионата у низших жи вотных, видимо, сходна с ролью
молочнокислого брожения в мышцах млекопитающих. Различные виды брожения, так же
как и фумаратное дыхание, могут доставлять некоторое количество энергии и
строго аэробным организ мам, что позволяет им переносить кратковременную аноксию. | 



