| Среда, 13.05.2026, 05:39 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В разделе материалов: 204
Показано материалов: 101-110 |
Страницы: « 1 2 ... 9 10 11 12 13 ... 20 21 » |
Железобактерия Thiobacillus ferrooxidans окисляет
двухвалентное железо до трехвалентного:
4Fe2+
+ 4Н+ + 02
-> 4Fe3+ + 2Н20
Эта бактерия
очень похожа на Т. thiooxidans, жизнеспособна при рН
среды до 2,5, однако энергию она получает не только за счет окисления
восстановленных соединений серы, но и за счет окисления ионов Fe2+.
Эта железобактерия обитает в кислых рудничных водах, содержащих сульфиды
различных металлов, в том числе пирит (FeS2). С несом ненностью
установлено, что ацидофильные железобактерии способ ны к хемоавтотрофному
образу жизни. |
Молекулярный
водород образуется в процессе анаэробного распада ор ганических веществ в
осадках водоемов и в анаэробных участках почвы. Многие бактерии способны к
использованию этого водорода. Значи тельная часть его подвергается окислению
теми бактериями, которые живут в сообществе с выделяющими Н2 организмами,
осуществляющи ми брожение. Окисление Н2 такими бактериями
сопровождается восста новлением сульфата до сульфида или С02 до
метана. По чти во всех группах бактерий, синтезирующих АТР путем окислитель
ного фосфорилирования в анаэробных условиях («анаэробное дыхание»), есть формы,
способные использовать молекулярный водород в качестве донора электронов. |
Бактерии,
окисляющие в аэробных условиях молекулярный водород с использованием кислорода
как конечного акцептора электронов, объе диняют в группу аэробных водородокисляющих
(водородных) бактерий. Все они способны как к автотрофной фиксации С02,
так и к использо ванию органических субстратов. Таким образом, водородные
бактерии являются факультативными хемолитоавтотрофами.
Некоторые
аэробные водородные бактерии могут окислять и СО, используя этот газ как
единственный донор электронов и единственный источник углерода. |
Большинство
организмов, способных расти в условиях, когда един ственным источником углерода
служит углекислота, фиксируют ее через рибулозобисфосфатный цикл
(цикл-Кальвина-Бассама). К таким организмам относятся аэробные
хемолитоавтотрофные бактерии, почти все фототрофные
бактерии, цианобактерии и
зеленые растения. Цикл Кальвина- Бассама определенно не
участвует в ассимиляции С02 у ме-танобразующих и ацетогенных
бактерий, хотя они тоже относятся к хе-молитоавтотрофным организмам.
Для
рибулозобисфосфатного цикла характерны два фермента, не участвующие в других
метаболических путях,-фосфорибулокиназа и рибулозобисфосфат-карбоксилаза.
Последний фермент представляет собой белок, в количественном отношении
преобладающий над всеми другими белками на нашей планете. Рибулозобисфосфатный
цикл -это восстановительный процесс, в котором С02 восстанавливается
до уров ня углеводов. В цикле могут быть выделены три участка: 1) реакция
карбоксилирования, 2) восстановление и 3) регенерация молекул, служа щих
акцепторами С02. |
Способность
использовать свет как источник энергии, необходимой для роста,
присуща двум группам бактерий, принципиально отличающимся друг от друга.
Пурпурные
и зеленые бактерии, объединяемые в порядок Rho-dospirillales1,
можно рассматривать как реликтовые организмы, дошед шие до нас из времен
начальной эволюции фотосинтеза. Они не в со стоянии использовать в качестве
донора водорода воду (как это делают зеленые растения); им требуются доноры с
более высокой степенью восстановления (H2S, H2 или
органические вещества). Поэтому фотосин тез у этих бактерий протекает без
выделения 02. В таких случаях гово рят об аноксигенном фотосинтезе.
Бактерии этой группы - типичные во дные организмы, распространенные как в
пресной, так и в морской воде. Их красная, оранжевая или зеленая окраска
обусловлена присут ствием бактериохлорофиллов и каротиноидов. |
Фототрофные
бактерии, осуществляющие аноксигенный фотосинтез, де лятся на две большие
группы: пурпурные бактерии (Rhodospirillales) и зеленые бактерии
(Chlorobiales). Представители этих двух порядков значительно различаются по
своим цитологическим и физиологическим признакам, а также по характерным для
них пигментам. |
Благодаря
фотосинтетическим пигментам достаточно густые суспензии фототрофных бактерий
имеют зеленую, сине-зеленую, пурпурно-фиоле товую, красную, коричневую или
розовую окраску. Цвет зависит от природы и количественного соотношения
пигментов. Отдельные пиг менты можно распознать даже по спектрам поглощения
интактных клеток (рис. 12.10). Хлорофиллы, например, ответственны за максимумы
поглощения в синей (< 450 нм) и в красной и инфракрасной (650-1100 нм)
областях спектра. Поглощение в области 400-550 нм обусловлено главным образом
каротиноидами, а у цианобактерий в области 550-650 нм-фикобилипротеинами. 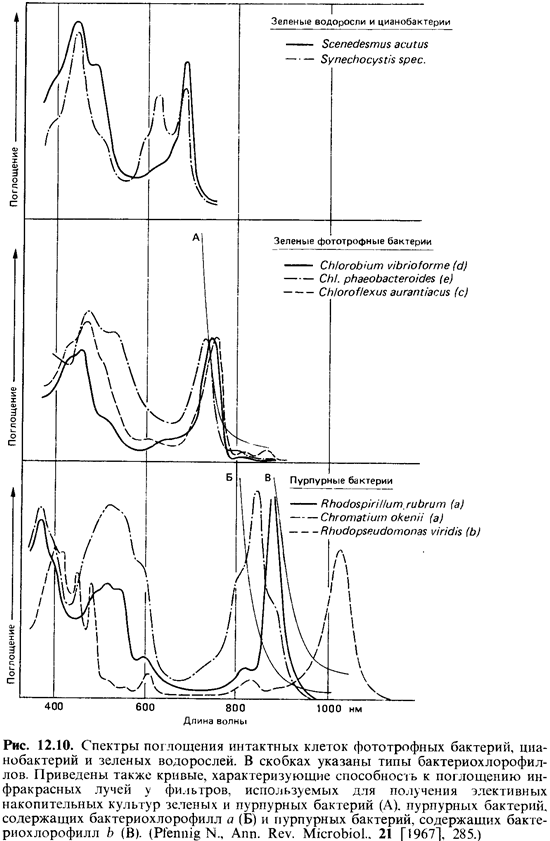
|
Исследование
обмена веществ у фототрофных бактерий сопряжено с ря дом трудностей. Эти
бактерии часто отличаются большой разносторон ностью. Например, многие несерные
пурпурные бактерии способны как к анаэробному росту на свету, так и к аэробному
росту в темноте; дру гие группы представлены строгими анаэробами и облигатными
фото-трофами. Многие виды используют в качестве донора электронов молекулярный
водород, а некоторые-сероводород или серу. Наблюдается ассимиляция С02 и
органических веществ. В анаэробных условиях в темноте за счет брожения может
получаться небольшое количество энергии, недостаточное, однако, для роста
большинства видов. Короче говоря, фототрофные бактерии относятся к организмам,
наиболее раз носторонним в отношении обмена веществ. Поэтому здесь мы можем
лишь глобально охарактеризовать их физиологические
возможности. |
Фототрофные
пурпурные и зеленые бактерии обитают в анаэробных зо нах многих водоемов, в
мелководных прудах, медленно текущих водах, в озерах и морских бухтах.
Пурпурные серобактерии нередко образуют налеты, окрашенные во всевозможные
оттенки красного цвета, от неж но-розового до темно-красного, на поверхности
ила или на каком-ни будь разлагающемся растительном материале. Иногда они
роятся над поверхностью ила, образуя слой толщиной около дециметра. Такое
«цветение» воды в мелких водоемах (рис. 12.12) вызывают в первую очередь
крупные представители пурпурных бактерий - Chromatium okeniu С.warmingii,
С. weissei и Thiospirillum jenense, однако
встречаются и мел кие представители Chromatiaceae и Chlorobiaceae. Бурное
размножение пурпурных серобактерий наблюдается и в таких мелководных прудах,
поверхность которых покрыта плотным слоем ряски (Lemna) или же листьями водяных
лилий. Этот своеобразный биологический фильтр по глощает те спектральные
компоненты света, которые могли бы исполь зоваться зелеными водорослями и
цианобактериями, но пропускает свет, поглощаемый бактериохлорофиллами и
темно-красными кароти-ноидами. Поэтому под покровом из ряски растут анаэробные
фото трофные бактерии, но не развиваются зеленые водоросли и цианобакте-рии
(рис. 12.12). Некоторые виды фототрофных серобактерий предста влены
в естественных местообитаниях почти чистой культурой. Всегда можно обнаружить и
Rhodospirillaceae, но лишь редко в столь массовом количестве. |
Метод отбора
анаэробных фототрофных бактерий основан на том (опи санном выше) наблюдении,
что пурпурные серные бактерии достигают иногда массового развития и в мелких
водоемах, если вся поверхность воды покрыта плотным слоем ряски. С помощью
фильтров, поглощаю щих коротковолновую часть спектра и пропускающих
инфракрасные лучи, используемые лишь отдельными группами фототрофных бакте рий,
можно создать селективные условия для роста как зеленых серо бактерий, так и
пурпурных бактерий, содержащих бактериохлорофилл а или b. |
|
 |  |


