| Пятница, 15.05.2026, 22:22 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В разделе материалов: 204
Показано материалов: 51-60 |
Страницы: « 1 2 ... 4 5 6 7 8 ... 20 21 » |
Глюкозо-6-фосфат
сначала, как это было описано выше для пентозофосфатного пути, дегидрируется до
6-фосфоглюконата. Под действием фосфоглюконатдегидратазы от него
отщепляется вода и образуется 2-кето-3-дезокси-6-фосфоглюконат (рис.
7.5). Кетодезоксифосфоглюконат расщепляется специфической альдолазой на пируват
и глицеральдегид-3-фосфат. Последний окисляется до пирувата, так же как и в
фруктозодисфосфатном пути. |
Пируват
занимает центральное положение в промежуточном метаболизме и может служить
предшественником разнообразных продуктов. Многие организмы окисляют большую
часть образующегося в ходе катаболизма пирувата до ацетилкофермента А.
Важнейшую роль у бактерий играют три реакции:
- (1) Пируват + СоА + NAD -> Ацетил-СоА + NADH2 +
СО2
- (2) Пируват + СоА + 2Fd -> Ацетил-СоА + 2FdH + СО2
- (3) Пируват + СоА -> Ацетил-СоА + Формиат
|
Цикл
трикарбоновых кислот (рис. 7.7) служит для окисления двухуглеродного соединения
ацетата до СО2 с отщеплением водорода. При участии трех
дегидрогеназ водород переносится на NAD(P), а под действием сукцинатдегидрогеназы
- непосредственно на хинон. Как правило, коферменты передают водород в
дыхательную цепь. |
В то время
как большинство анаэробных организмов способно синтезировать АТР только путем
фосфорилирования на уровне субстрата, аэробы могут осуществлять несравненно
более эффективную регенерацию АТР. Они обладают особым аппаратом: дыхательной
(электрон-транс портной) цепью и ферментом ATР-синтазой;
обе системы у прокариот находятся в плазматической мембране, а у эукариот – во
внутренней мембране митохондрий. Ведущие свое происхождение от субстратов
восстановительные эквиваленты (Н или электроны) в этих мембранах по ступают в
дыхательную цепь, и электроны переносятся на О2 (или другие
терминальные акцепторы электронов). В дыхательной цепи происходят реакции,
представляющие собой биохимический аналог сгорания водорода. От химического
горения молекулярного водорода они отличаются тем, что значительная часть
свободной энергии переводится при этом в биологически доступную форму, т. е. в
АТР, и лишь небольшая доля рассеивается в виде тепла. |
Транспорт
водорода и транспорт электронов - процессы эквивалентные. Дыхательная цепь
может рассматриваться как цепь переноса электронов. Компоненты дыхательной цепи
переходят попеременно из окисленного состояния в восстановленное и обратно, т.
е. ведут себя как типичные окислительно-восстановительные катализаторы. Они
обладают окислительно-восстановительным потенциалом, который может быть измерен
непосредствен но (у цитохромов) или косвенно (у NAD, FAD). |
По
окислительно-восстановительному потенциалу компоненты дыхательной цепи можно
расположить в ряд, который начинается с NAD (наиболее отрицательный потенциал)
и оканчивается цитохромоксидазой и кислородом (рис. 7.11). Хиноны и цитохромы
служат вспомогательными субстратами. Эти компоненты восстанавливаются
водородом, доставляемым различными донорами. Получаемый через NAD водород с
помощью NADH-дегидрогеназы переносится на хинон; аналогичным образом на хинон
переносится водород от сукцината (через сукцинатдегидрогеназу) и водород,
полученный при дегидрировании жирных кислот (через другие специфические
дегидрогеназы). Хинон служит сборным резервуаром водорода из субстратов в дыхательной
цепи. Некоторые из ферментов, участвующих в переносе водорода, содержат FMN или
FAD, а также железосерные белки. |
Дыхательная
цепь ингибируется или блокируется различными клеточными ядами. Амитал, ротенон
и пиерицидин А подавляют активность NADH-дегидрогеназы; антимицин А блокирует
перенос электронов между цитохромами b и с. Цианид и окись углерода
ингибируют только цитохромоксидазу; в цитохроме с ион железа,
по-видимому, так спрятан в глубине белка, что не взаимодействует ни с CN
", ни с СО. Специфическое действие этих ядов и изменения характерных
спектров поглощения компонентов дыхательной цепи послужили индикаторами, с
помощью которых была изучена эта цепь. |
Рассмотрение окислительно
- восстановительных потенциалов (табл. 7.4) показывает, что в дыхательной цепи
имеются только три этапа окисления, на которых освобождается по меньшей мере
столько энергии, сколько содержится в одной «высокоэнергетической» связи. При
переносе 2 [Н] от NADH2 на кислород только три электронных
перехода могут быть сопряжены с фосфорилированием ADP в АТР, так что в лучшем
случае лишь три молекулы фосфата могут быть включены в органическое соединение.
Эту связь окисления с фосфорилированием обычно выражают в виде коэффициента Р/О
(число молекул АТР, образующихся на 1 атом затраченного кислорода). Для
митохондрий животных, используя в качестве доноров водорода изоцитрат или малат
(переносящие свой водород на NAD), можно экспериментально получить коэффициент
Р/О, равный 3; для сукцината, от которого водород может быть включен в цепь
дыхания только на уровне флавопротеинов, коэффициент Р/О составляет лишь 2. 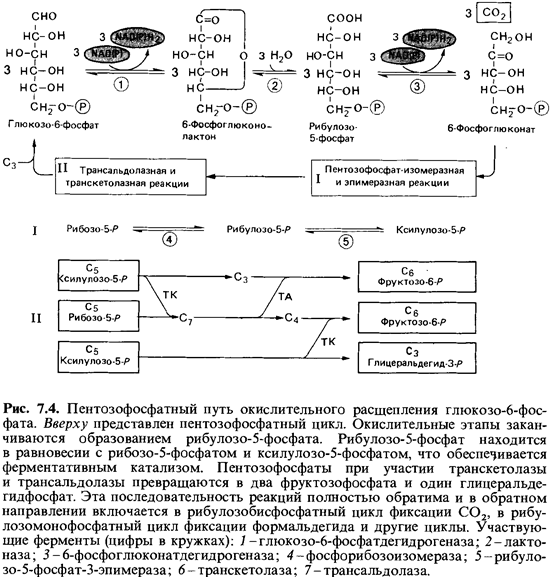
|
Регенерация
АТР при фосфорилировании в дыхательной цепи и фотосинтетическом фосфорилировании
протекает в мембранах. АТР-синтаза, так же как и компоненты дыхательной цепи,
является составной частью мембраны. Каким образом происходящий в дыхательной
цепи перенос водорода и электронов сопряжен с синтезом АТР, до конца еще не
выяснено. Однако многочисленные эксперименты показали, что регенерация АТР
происходит только в пространствах, окруженных со всех сторон мембранами, - в
пузырьках, или везикулах. Процессы переноса водорода и электронов теснейшим
образом сопряжены с перемещением протонов, а этот процесс в свою очередь
необходим для регенерации АТР. |
Добавляя молекулярный
кислород к суспензии аэробных бактерий или митохондрий, находившихся перед тем
в анаэробных условиях, можно заметить снижение рН среды. Это позволяет
заключить, что во время дыхания из бактериальных клеток и митохондрий выводятся
протоны (рис. 7.12, А и Б). Если из мембран бактерий или
митохондрий приготовить пузырьки, у которых прежняя внутренняя сторона обращена
наружу (вывернутые везикулы), то при дыхании будет наблюдаться «обращенный»
перенос протонов, приводящий к подщелачиванию суспензионной среды (рис. 7.12,
В). |
|
 |  |


