| Среда, 13.05.2026, 04:01 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В разделе материалов: 204
Показано материалов: 131-140 |
Страницы: « 1 2 ... 12 13 14 15 16 ... 20 21 » |
Среди
углеводородов метан занимает особое положение. Его исполь зуют и окисляют
бактерии, не способные расщеплять углеводороды с длинной цепью. Такого рода
бактерии следует рассматривать не как микроорганизмы, окисляющие углеводороды,
а скорее как группу бак терий, специализировавшихся на использовании
Ci-соединений. Поэто му бактерии, окисляющие метан, относят вместе со всеми
бактериями (и дрожжами), способными использовать метанол, метилированные амины,
диметиловый эфир, формальдегид и формиат, к метило-трофным организмам. В
накопительных культурах с метаном как единственным источником углерода и
энергии развиваются бактерии, отно сящиеся к различным родам:Methylomonas, Methylococcus, Methylosinus. Некоторые
из них растут только на средах, содержащих метан, метанол или диметиловый эфир,
и не могут использовать сахара, органические кислоты или иные спирты (кроме
метанола). |
В то время
как в накопительных культурах, содержащих помимо С02 и О 2 только
чистый метан, развивается главным образом Methylomonas methanica, в
культурах на природном газе, в состав которого наряду с метаном входит и этан,
растут одни только окислители этана. К окис лению этана способно гораздо
большее число видов, чем к окислению метана. Большинство микроорганизмов,
окисляющих этан, принадле жит к родам Mycobacterium, Flavobacterium и Nocardia. Некоторые
из бактерий, использующих этан, могут также окислять газообразный во дород. Еще
большее число видов бактерий развивается в накопи тельных культурах за счет
использования пропана. Были выделены так же бактерии, способные окислять бутан (Mycobacterium и Pseudomonas). |
Алканы с
длинной цепью используются очень многими бактериями. Ре шающее значение при
этом имеет длина цепи: по мере удлинения цепи парафинов растет число видов,
способных использовать эти соединения, а также активность их использования. В
их разложении участвуют ми-кобактерии, нокардии и коринебактерии. Долгое время
способность бактерий расти на средах с углеводородами рассматривалась как
своего рода курьез. Интерес к микроорганизмам, окисляющим углеводороды, возник
в связи с двумя наблюдениями. В 1950
г. в Институте бро дильных производств в Берлине из
накопительных культур, содержавших в качестве источника энергии различные
фракции углеводородов, были выделены два вида дрожжей - Candida lipolytica и Candida tropicalis. Оказалось,
что С. lipolytica использует все высшие гомологи углеводо
родов, начиная с С15-соединений. Вслед за этим было установлено, что
большинство видов Candida окисляет углеводороды. Проведенная
про верка штаммов из коллекции дрожжей показала, что способность ис пользовать
углеводороды присуща очень многим из них. Использова ние углеводородов
протекает с необычайно высоким выходом полезных продуктов (У). Если при
использовании углеводов коэффициент У со ставляет около 0,5, то для
углеводородов он колеблется в пределах от 0,7 до 1,0. |
Растения
синтезируют много соединений, содержащих ароматические кольца. Из них в
количественном отношении преобладает лигнин, со ставляющий 20% (по весу)
древесины. Способность расщеплять такие соединения с разрывом ароматического
кольца обладают многие бакте рии и грибы. Некоторые псевдомонады растут на
среде, содержащей бензоат, быстрее, чем на среде с сахарами. Для быстрого
расщепления ароматических веществ необходимо присутствие молекулярного кисло
рода. Пути такого расщепления мы рассмотрим в этом разделе^ Соглас но новейшим
данным, ароматические соединения могут подвергаться также анаэробному
разложению, но мы не будем описывать его мета болические механизмы. |
Азот
органических соединений - это прежде всего белковый азот. Подобно другим
высокомолекулярным соединениям, белки сначала рас щепляются внеклеточными
протеазами на фрагменты, способные проникнуть в клетку, - полипептиды,
олигопептиды и отчасти аминокис лоты. Пептиды поступают в клетки и
гидролизуются внутриклеточны ми протеазами до аминокислот. Последние либо
используются клеткой как таковые для синтеза белка, либо подвергаются
превращениям, в ре зультате которых они в конечном счете дезаминируются и после
этого вовлекаются в промежуточный обмен (рис. 14.14). 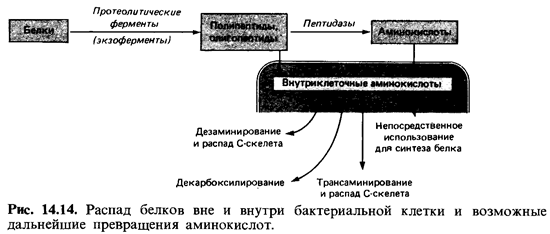
|
Всякое живое
существо по большинству своих признаков сходно со своими предками. Сохранение
специфических свойств, т.е. постоянство признаков в ряду поколений, называют наследственностью.Изучением
передачи признаков и закономерностей их наследования занимается генетика. Каждому
признаку в качестве носителя информации соответ ствует определенный ген. Еще
во времена классической генетики иссле дователи пришли к выводу, что гены
находятся в клеточном ядре. Тогда же было установлено, что они должны
располагаться в линейном порядке. Долгое время считали, что наследственная
информация связа на с белковыми компонентами нуклеоплазмы. Лишь после успешных
экспериментов по передаче наследственных признаков с помощью ДНК генетики
пришли к убеждению, что именно ДНК, входящая в состав хромосом у всех
организмов, служит материальным носителем наследственной информации. Сначала
на насекомых, а затем на микроорганизмах было показано, что проявление
признаков зависит от активности ферментов. У микроорганизмов ферменты можно
было свя зать с конкретными признаками, поддающимися точному биохимиче скому
определению. Гипотеза один ген-один фермент гласит, что опре
деленный ген содержит информацию, необходимую для синтеза определенного
фермента (позднее была принята более точная формули ровка: каждый структурный
ген кодирует определенную полипептид ную цепь). Изменение гена вследствие
мутации приводит либо к утрате фермента, либо к изменению его свойств, а тем
самым и к изменению признака. Гены выявляются только благодаря мутациям.
Генетический анализ основан прежде всего на изучении различий в признаках,
опреде ляемых альтернативными формами (аллелями) того или
иного гена. По этому исследование различных генетических проблем ведется на
мутан тах. |
Репликация
ДНК. У бактерий, так же как и у высших организмов, носи телем
генетической информации служит ДНК. Рассматривая структуру клетки, мы уже
говорили о том, что бактериальная ДНК представляет собой двойную спираль,
замкнутую в кольцо. Сразу же возникает во прос: как сохраняется наследственная
информация при росте и размно жении клеток? Перед их делением происходит идентичная
редупликация,или репликация, генов. Этот процесс можно
удовлетворительно объяс нить, исходя из модели структуры ДНК, предложенной
Уотсоном и Криком, и из механизма удвоения ДНК, теперь уже известного. Две цепи двойной спирали ДНК комплементарны друг другу. На каждой цепи из
структурных элементов ДНК-дезоксирибонукле-озидтрифосфатов-синтезируется новая
цепь; при этом с каждым из ос нований спаривается комплементарное ему
основание, так что каждая из двух новых цепей опять-таки будет комплементарна
родительской цепи. Обе новые двойные спирали состоят из одной родительской и
одной вновь синтезированной цепи. Эта точная репликация ДНК га рантирует
сохранение генетической информации. |
Представление
о том, что и у микроорганизмов возможны скачко образные изменения
наследственных признаков-мутации,-утвержда лось лишь с трудом. До разработки
метода чистой культуры многие ученые (Нэгели, Цопф) думали, что у бактерий
морфология и физиоло гические свойства чрезвычайно изменчивы. Считалось, что
большое чис ло бактерий, встречающихся в природе, представляют собой разные
стадии жизненного цикла небольшого количества видов (плеоморфизм). Возражая
против этого на основании результатов, полученных с по мощью
усовершенствованных методов и чистых культур, другие ученые выступили в пользу
теории мономорфизма, согласно которой бактерии можно различать
и классифицировать, исходя из постоянства их морфо логических и физиологических
признаков. Необходимо было научиться различать и у бактерий генотип и фенотип. Генотипом называют
сово купность наследственных задатков клетки; ему противопоставляют фенотип - совокупность наблюдаемых признаков. Фенотипическое проявле ние одного и
того же генотипа может быть различным в зависимости от условий среды. |
Спонтанные мутации В популяции бактерий без всякого экспериментального вмешательства регулярно возникают мутации; такие мутации называют спонтанными мутациями, а клетки, в которых они возникли,-спонтанными мутантами. Мутагенное действие аналогов оснований ДНК (см. ниже) указывает на возможные причины спонтанных мутаций: вероятно, речь идет о случайных ошибках при включении нуклеотидов во время репликации ДНК-ошибках, вызванных таутомерным перемещением электронов в основании. Тимин, например, обычно находится в оксо-форме, в кото рой он образует водородные связи с аденином. Но если тимин во время спаривания оснований при репликации ДНК переходит в енольную форму, то он спаривается с гуанином. В результате в новой молекуле ДНК на том месте, где раньше находилась пара А-Т, появляется пара G-C. |
|
На обычных твердых средах лишь немногие мутации можно непосред ственно обнаружить по изменению пигмента, измененному росту коло ний или иным признакам. Некоторые мутантные признаки выявляются при добавлении индикаторов или красителей. Для идентификации му тантов, отличающихся от родительских клеток пониженными или повы шенными требованиями к питанию, приходится сравнивать рост тех и других на двух средах. Если, например, мутант утратил способность к синтезу лейцина, которой обладали клетки родительского (дикого) ти па, то он будет расти только на той среде, к которой добавлена эта аминокислота. Мы называем такого мутанта ауксотрофным по лейцину, т.е. нуждающимся в лейцине (1еи~), а также дефектным по лейцину, противопоставляя ему прототрофный дикий тип (leu +). Если в клеточной суспензии присутствуют одновременно и мутантные клетки leu ~, и про-тотрофные клетки дикого типа, то эти два типа можно различить по росту на двух разных средах. Метод, обычно применяемый для выявле ния таких дефектных. |
|
 |  |


