Среди
углеводородов метан занимает особое положение. Его исполь зуют и окисляют
бактерии, не способные расщеплять углеводороды с длинной цепью. Такого рода
бактерии следует рассматривать не как микроорганизмы, окисляющие углеводороды,
а скорее как группу бак терий, специализировавшихся на использовании
Ci-соединений. Поэто му бактерии, окисляющие метан, относят вместе со всеми
бактериями (и дрожжами), способными использовать метанол, метилированные амины,
диметиловый эфир, формальдегид и формиат, к метило-трофным организмам. В
накопительных культурах с метаном как единственным источником углерода и
энергии развиваются бактерии, отно сящиеся к различным родам:Methylomonas, Methylococcus, Methylosinus. Некоторые
из них растут только на средах, содержащих метан, метанол или диметиловый эфир,
и не могут использовать сахара, органические кислоты или иные спирты (кроме
метанола).
Получение
энергии. Восстановительные эквиваленты для получения энергии могут
создаваться в результате окисления метана через этапы образования метанола,
формальдегида и формиата до С02. Окисление метана в метанол
сопровождается включением в молекулу одного ато ма кислорода; эту реакцию
катализирует метаноксигеназа.
Синтез
клеточного вещества. Вещество клеток синтезируется, как правило, из
формальдегида-промежуточного продукта окисления мета на. Этот синтез может
осуществляться разными путями, из которых на иболее изучены рибулозомонофосфатный
цикл фиксации формальдегида и сериновый путь.
В
рибулозомонофосфатном цикле формальдегид (продукт окисления метана)
.превращается в результате альдольной конденсации с рибуло зо-5-фосфатом
в арабино-3-гексулозо-6-фосфат; последний в ходе изо-меразной реакции
превращается в фруктозо-6-фосфат (рис. 14.6). Из фруктозо-6-фосфата снова
(подобно тому, как это происходит при фиксации С02 в
рибулозобисфосфатном цикле) образуется пентозофосфат. Цикл не включает реакцию,
катализируемую седогептулозо-1,7-фосфа-тазой. 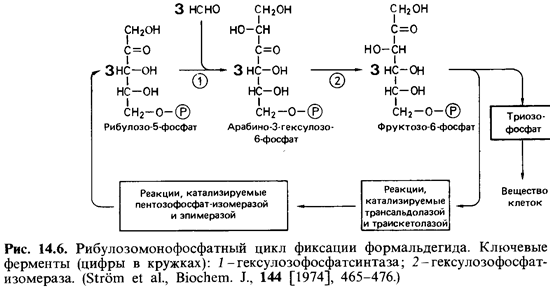
Серииовый
путь (рис. 14.7) был изучен на Pseudomonas MA и Нуphomicrobium X. Вероятно, у других микроорганизмов
имеются модифи кации этого пути. Акцептором формальдегида служит глицин.
Следует ожидать, что у организмов, ассимилирующих С1-соединения с помощью
этого цикла, удается обычно обнаружить гидроксипируватре-дуктазу,
малаттиокиназу, малил-СоА-лиазу и изоцитрат-лиазу. 
Дрожжи способны использовать только метанол,
но не метан. У ря да дрожжей (Candida boidinii, Hansenulapolymorpha) включение
метанола в вещество клеток происходит тоже через формальдегид, однако через
другой пентозофосфат, а именно ксилулозо-5-фосфат. В этом ксилулозо-монофосфатном
цикле фиксации формальдегида последний вместе с кси-лулозо-5-фосфатом
превращается с помощью специальной транскето-лазы в глицеральдегидфосфат и
дигидроксиацетон. Оба продукта (дигидроксиацетон - после фосфорилирования
триокиназой) поступают на пути синтеза.
Использование метанола бактериями начинается
при участии метанолдеги-дрогеназы. В этом ферменте была обнаружена ранее
неизвестная простетическая группа - метоксатин, или пирролохинолинхинон. В
настоящее время известно, что метоксатин входит в состав многих бактериальных
мембраносвязанных алкогольдегидрогеназ. | 



