Алканы с
длинной цепью используются очень многими бактериями. Ре шающее значение при
этом имеет длина цепи: по мере удлинения цепи парафинов растет число видов,
способных использовать эти соединения, а также активность их использования. В
их разложении участвуют ми-кобактерии, нокардии и коринебактерии. Долгое время
способность бактерий расти на средах с углеводородами рассматривалась как
своего рода курьез. Интерес к микроорганизмам, окисляющим углеводороды, возник
в связи с двумя наблюдениями. В 1950
г. в Институте бро дильных производств в Берлине из
накопительных культур, содержавших в качестве источника энергии различные
фракции углеводородов, были выделены два вида дрожжей - Candida lipolytica и Candida tropicalis. Оказалось,
что С. lipolytica использует все высшие гомологи углеводо
родов, начиная с С15-соединений. Вслед за этим было установлено, что
большинство видов Candida окисляет углеводороды. Проведенная
про верка штаммов из коллекции дрожжей показала, что способность ис пользовать
углеводороды присуща очень многим из них. Использова ние углеводородов
протекает с необычайно высоким выходом полезных продуктов (У). Если при
использовании углеводов коэффициент У со ставляет около 0,5, то для
углеводородов он колеблется в пределах от 0,7 до 1,0.
Механизм. Многие
псевдомонады окисляют углеводороды настоль ко полно, что накопления
промежуточных продуктов не происходит. Только у Acinetobacter calcoaceticus продукты
окисления выделяются в среду, а у Nocardia накапливаются в
клетке. Характер продуктов зави сит от природы субстрата. Если A. calcoaceticus растет
на среде с гекса-деканом, то из культуральной жидкости можно выделить цетилпальми-тат.
Цетилпальмитат представляет собой эфир пальмитиновой кислоты и цетилового
спирта (гексадеканола)-двух продуктов окисления гекса декана. На приводимой
схеме (рис. 14.8) показано, что расщепление па рафина начинается с окисления
концевого углерода. 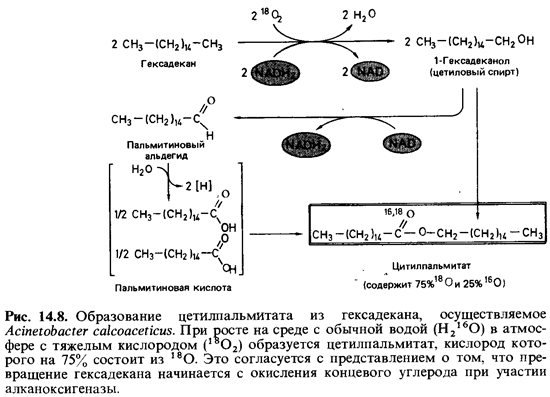
В первичном
воздействии на углеводородную цепь участвует кисло род. Без молекулярного
кислорода парафин не окисляется. Окисление катализирует монооксигеназа
(алканоксигеназа):
Парафин + 02 +
NADH2 -* Алифатический спирт + NAD +. Н20
Дальнейшее
окисление парафина протекает по пути, который известен как р-окисление жирных
кислот с длинной цепью (рис. 14.9). Если псев домонады, окисляющие гептан,
поместить в условия пониженного парциального давления кислорода, то в
культуральной жидкости будут на капливаться С3-, С5- и С7-жирные
кислоты. Вырастив клетки бактерий в присутствии гексана, а затем инкубируя их с
гептаном, можно наблюдать накопление пропионовой кислоты. Это, видимо,
объясняется тем, что клетки, выросшие в присутствии гексана, не содержат
ферментов, необходимых для окисления
пропионата через метилмалонил-СоА. 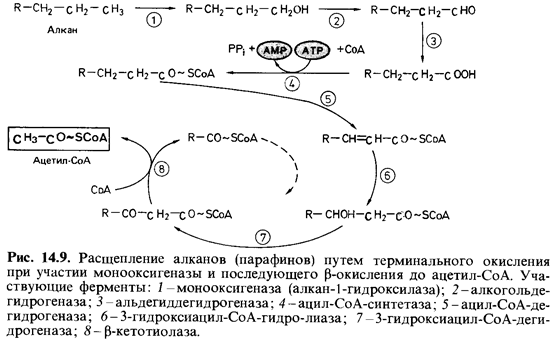
| 



