Первичные
процессы фотосинтеза протекают в тилакоидах - плоских
замкнутых мембранных пузырьках, содержащихся в клетках цианобак терий и в
хлоропластах водорослей и высших растений.
Тилакоидные
мембраны и светособирающие пигменты (пигменты антенн). Тилакоидная
мембрана содержит в себе пигментные молекулы (хлорофилл а, хлорофилл b и каротиноиды),
переносчики электронов и ферменты. Подавляющее большинство молекул хлорофилла (
> 99,5%), а также дополнительные пигменты (каротиноиды, фикобили-протеины)
ответственны за поглощение света и распределение энергии; они образуют систему
антенны. Лишь незначительная часть хлорофил ла авыполняет роль
фотохимического реакционного центра, в котором протекает собственно
фотохимическая окислительно-восстановительная реакция. Пигменты антенн
(светособирающие пигменты) улавливают свет и передают энергию хлорофиллу
реакционного центра (Каротиноид -> Каротиноид*; Хлорофилл + Каротиноид*
-> Хлоро филл* + Каротиноид). Каротиноиды выполняют также защитную функцию:
при очень ярком солнечном освещении они отдают избыточ ную энергию в окружающую
среду и тем самым защищают молекулы хлорофилла от фотоокисления. Система
светособирающих пигментов и реакционный центр объединены в так называемую
фотосинтетиче скую единицу.
Фотореакции. Фотореакции
относятся к первичным процессам любо го фотосинтеза. Местом, где протекают эти
фотохимические окисли тельно-восстановительные реакции, являются реакционные
центры. Реакционный центр состоит из ряда компонентов, наиболее
важные из которых - первичный донор электронов (особый комплекс из хлорофил ла
и белка) и первичный акцептор электронов. Эти два компонента представляют собой
окислительно-восстановительные системы. Систе ма донора (Р/Р+)
обладает положительным, а система акцептора (Х/Х ~)- отрицательным потенциалом.
Под воздействием энергии света происходит перенос одного электрона:
Восстановлен-
Окисленный hv
Окисленный Восстановлен-
+
-»
+
ныи
донор
акцептор
донор ный акцептор
Вместо этого
можно написать
Следовательно,
первую фотореакцию можно представить следующим образом:
Аналогичным
образом можно представить и фотореакцию у пурпурных бактерий:
Таким
образом, в результате фотореакции донор теряет один элек трон - возникает
«дырка» (электронный дефект). Такие «дырки» должны заполняться электронами,
которые могут поступать сюда по одному из двух путей-по пути нециклического или циклического
переноса электронов. При нециклическом переносе электроны поступают от
экзогенного внешнего донора: в случае второй фотореакции-от молекул воды, в
случае первой реакции-из электрон-транспортной цепи, связывающей обе
фотосистемы между собой. При циклическом переносе электроны возвращаются от
восстановленного акцептора (X ~) к окисленному до нору. Фотохимическая
окислительно-восстановительная реакция, в ходе которой Р окисляется, а X
восстанавливается, представлена на следую щей схеме:
Циклический
перенос электронов приводит к изменению заряда мем браны, а нециклический
перенос-кроме того, и к восстановлению NADP.
Две
фотореакции в двух пигментных системах (фотосистемах). При
оксигенном
фотосинтезе работают две пигментные системы, вклю ченные последовательно (рис.
12.14). Пигментную систему, возбуждае мую более длинноволновым светом (к
< 730 нм), называют фотосисте мой I, а возбуждаемую более
коротковолновым светом (X < < 700 нм) -
фотосистемой II. Фотохимически активный реакционный центр фотосистемы I
содержит Хл а.\ (Р700), играющий роль первичного донора электронов
в первой фотореакции. Световая энергия, поглощае мая светособирающими
пигментами фотосистемы I, передается в реак ционный центр и переводит в
возбужденное состояние Хл а\. Это приво дит к окислению Хла;,
т.е. к отдаче им одного электрона. Хл а\ превращается при этом
в Хл а\ . Другими словами, в результате отдачи электрона в
реакционном центре образуется «дырка», или «электронная вакансия». Эта «дырка»
тотчас же заполняется другим электроном, по ступающим по специальному
электрон-транспортному пути. Акцепто ром отданного электрона, по-видимому,
служит железосерный белок («X»). Он обладает еще более отрицательным
окислительно-восстанови тельным потенциалом, чем - 420 мВ, возможно - 530 мВ.
Этот акцеп тор в свою очередь отдает электрон ферредоксину, а с восстановленно
го ферредоксина восстановительная сила может передаваться на NADP или другие
акцепторы. Наряду с этим возможен и циклический перенос электрона, при котором
электрон от «X» передается через пластохинон, цитохромы и пластоцианин обратно
к хлорофиллу а\ реакционного центра. 
Реакционный
центр фотосистемы II содержит Хл ац (Р680), который служит
первичным донором электронов во второй фотореакции. Полу чив энергию,
поглощенную светособирающими пигментами фотоси стемы II, этот хлорофилл
переходит в возбужденное состояние. Возбуж дение Хл а\\ ведет
к эмиссии одного электрона, являющегося слабым восстановителем (Е'0 ~
О В). Этот электрон принимает молекула особо го пластохинона (Х320), который
при этом восстанавливается до семи-хинона. Донором электронов для фотосистемы
II служит вода. «Дыр ка», образовавшаяся в Хл а\\ в результате
потери электрона, заполняется одним из электронов, освобождающихся при
образовании 02 из воды (2Н20 -> 02 +
4Н + + 4е ~). Разложение воды происходит при участии
марганца.
Две описанные
выше пигментные системы связаны между собой электрон-транспортной цепью, важным
звеном которой является пла-стохинон. Подобно убихинону в дыхательной цепи,
пластохинон в фотосинтетической электрон-транспортной цепи находится в большом
из бытке и выполняет функцию накопителя (депо) электронов. Этот накопитель
может вмещать не менее 10 электронов (на 1 молекулу Хл ац), поступающих от
Х320. Окисление пластохинона осуществляет фо тосистема I, т.е. электроны
«накопителя» расходуются на заполнение «дырок» в Хл а\ . От
пластохинона электроны передаются цитохрому / (мембраносвязанному цитохрому
типа с), затем пластоцианину (раст воримый медьсодержащий белок) и, наконец,
хлорофиллу а\ . Таким образом, пластохинон выполняет важную
функцию накопления и даль нейшей передачи электронов, поступающих из нескольких
(как минимум десяти) электрон-транспортных цепей.
Основные пути
транспорта электронов в ходе первичных процессов фотосинтеза показаны на рис.
12.14. Это известная Z-схема- результат исследований, в
которых использовались методы импульсной спектро-фотометрии, а также
искусственные доноры и акцепторы электронов и специфические ингибиторы. Она
дает представление об окислительно-восстановительных потенциалах пигментов и
переносчиков электронов и о последовательности их окисления и восстановления,
но ничего не говорит о локализации этих компонентов в мембране.
Локализация
пигментов и переносчиков электронов в мембране. Неко торые сведения по
этому вопросу были получены при исследовании функций тилакоидов в присутствии
антител, а также липофильных или гидрофильных искусственных
окислительно-восстановительных систем. Иммунные антитела, полученные против
отдельных очищенных компо нентов фотосинтетической электрон-транспортной
системы, не в состоя нии проникать сквозь мембрану и реагируют поэтому только с
теми компонентами, которые расположены на наружной поверхности тила коидов.
Например, антитела к ферредоксину или к ферредоксин-NADP-редуктазе эффективно
подавляют функцию фотосистемы I; значит, эти два компонента находятся на
внешней поверхности тилакоидов. В то же время поверхность, на которой
расположены доноры электронов, недо ступна для антител. Хотя данные о
структурной организации фотосистемы II еще весьма противоречивы, уже можно
привести гипотетиче скую схему (рис. 12.15). Эту схемупространственной ориентации
направленного фотосинтетического транспорта электронов внутри тилакоидной
мембраны следует отличать от схемы на рис. 12.14, где компо ненты расположены
по вертикали в соответствии с их окислительно-восстановительными потенциалами.
Схема на рис. 12.15 наглядно показывает, что электроны, освобождающиеся в
результате расщепле ния воды, переносятся из внутреннего пространства
тилакоидов в строму. 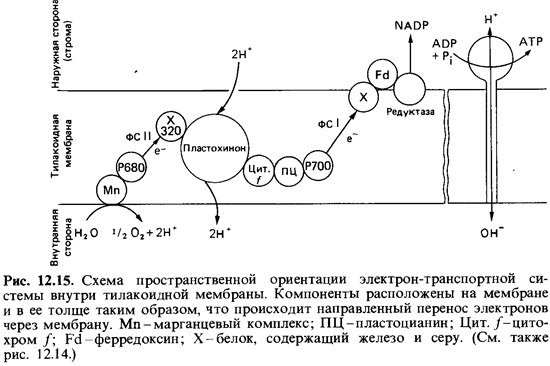
Направленный
транспорт электронов и создание протонного градиента. Представления о
локализации компонентов фотосинтетической элек трон-транспортной цепи
согласуются с данными физиологических на блюдений и измерений. Если суспензию
тилакоидов или разрушенных хлоропластов подвергнуть воздействию света, то рН
суспензионной среды возрастает, а после выключения света снова понижается. Свет
вызывает передвижение протонов внутрь тилакоидов (рис. 12.16). Таким образом,
световая энергия может использоваться для создания градиен та протонов на
тилакоидной мембране. Еще раньше было известно, что повышение рН в суспензии
тилакоидов с 4 до 8 (в темноте) приводит к синтезу АТР. Подобного рода
эксперименты легли в основу гипотезы о хемиосмотическом механизме
преобразования энергии. Более де тальные исследования показали, что перенос
одного электрона через обе фотосистемы сопровождается поступлением двух
протонов во вну треннее пространство тилакоида. Как сейчас полагают, две фотоси
стемы вместе со связывающей их электрон-транспортной цепью обеспе чивают
направленный поток электронов от воды (с внутренней стороны тилакоидной
мембраны) к NADP (с внешней стороны). Таким образом, фотореакции ведут к
восстановлению NADP и образованию заряда на мембране. Иными словами, световые
реакции выступают в роли про тонного насоса, который работает
за счет энергии света и создает поло жительный заряд внутри тилакоида; в
результате мембрана аккумули рует энергию в форме протонного потенциала, и эта
энергия используется для синтеза АТР. Протонный потенциал связывает фото
синтетический транспорт электронов с фосфорилированием таким же образом, как он
связывает с фосфорилированием транспорт электронов при дыхании. 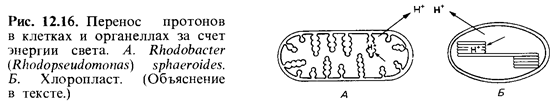
| 



