Фотосинтетический
транспорт электронов у анаэробных фототрофных бактерий во многих отношениях
отличается от только что описанного. В аноксигенном фотосинтезе участвует
только одна световая реакция; она поддерживает циклический транспорт
электронов. Электроны, поки дающие цикл для восстановления NAD, не являются
продуктом разло жения воды. Фотосинтез зависит от наличия в среде
восстановленных субстратов и не сопровождается выделением 02.
Собственно фотореак ция хотя и аналогична первой фотореакции у зеленых
растений, однако у некоторых бактерий она приводит, вероятно, лишь к созданию
про тонного потенциала и тем самым к запасанию энергии (АТР), но не к
восстановлению NAD. Таким образом, нециклический перенос элек тронов (от донора
электронов к пиридиннуклеотиду) здесь отсутствует. По-видимому, NADH2 образуется
в результате какой-то темновой реак ции в ходе обратного транспорта электронов,
протекающего с затратой энергии.
Следует,
однако, заметить, что у фототрофных бактерий между от дельными группами
существуют гораздо большие различия в составе пигментов и механизмах
фотосинтеза, чем у зеленых растений. В после дующем изложении мы вначале не
будем касаться зеленых бактерий.
Фотореакция
у пурпурных бактерий. Как уже говорилось, у пур пурных бактерий
пигменты и компоненты электрон-транспортной си стемы тоже находятся в
мембранах. Пигментный комплекс фотохими ческого реакционного центра удается
отделить от пигментов антенны.
Энергия,
поглощенная пигментами антенны (бактериохлорофиллом и каротиноидами) передается
реакционным центрам. Изолированные реакционные центры состоят из белкового
комплекса, содержащего Бхл а, бактериофеофитин, каротиноиды,
убихинон и железосерный белок (FeS-белок). Пигмент реакционного центра
обозначают Р870~по длине волны, при которой максимально снижается поглощение
под действием света. На свету Р870 окисляется в Р870+.
Окислительно-восстанови тельный потенциал этого донора электронов лежит между +
450 и + 490 мВ. Первичным акцептором электронов, вероятно, служит ком плекс
убихинона с FeS-белком. Окислительно-восстановительный потен циал этого
комплекса должен быть близок к -100 мВ. Поэтому кажется маловероятным, что
электроны, возбуждаемые при световой реакции у пурпурных бактерий, способны
восстанавливать NAD. Ско рее, они возвращаются через убихинон, цитохромы Ь и
с2 и, возможно, FeS-белки назад к Р870 + (рис.
12.17). Необходимые же для восстановле ния NAD электроны, видимо, покидают путь
циклического транспорта. Они переносятся на NAD в результате обратного
транспорта, проте кающего с затратой АТР. Это существенное отличие
от в остальном аналогичной первой фотореакции при оксигенном фотосинтезе. Для
по полнения цикла электронами пурпурные бактерии нуждаются во внеш них донорах
электронов. Пурпурные серобактерии могут использовать с этой целью сероводород,
серу или тиосульфат; органические соедине ния (малат, сукцинат и др.) и
молекулярный водород служат донорами электронов для обеих групп пурпурных
бактерий.
Как показали
многочисленные эксперименты, и у пурпурных бакте рий фотосинтетический перенос
электронов приводит к созданию протонного градиента. Интактные клетки реагируют
на воздействие света выделением в среду протонов, приводящим к закислению
среды. В сус пензии пузырьков из фотосинтетических мембран (хроматофоров) свет
вызывает перенос протонов, направленный внутрь. Таким образом, мембраны
хроматофоров и тилакоидов имеют такую же полярность, как и субмитохондриальные
пузырьки. Это будет понятно, если учесть, что все эти мембраны образуются путем
впячивания внутрь и разраста ния плазматической мембраны или же внутренней
мембраны хлоропла ста. Хотя точная локализация отдельных компонентов в мембране
еще не установлена, можно думать, что переносчики водорода и электронов
расположены и в мембране анаэробных фототрофных бактерий таким образом, что
происходит разделение зарядов. В хроматофорах элек троны транспортируются
наружу, а протоны - внутрь. Создающийся протонный потенциал и служит движущей
силой фотосинтетического фосфорилирования.
Фотореакция
у зеленых бактерий. Механизмы фотореакции у зеленых бактерий еще не
полностью выяснены. Есть указания на то, что пер вичный акцептор электронов,
участвующий в световой реакции, у зеленых серобактерий обладает потенциалом
около - 500 мВ (у пур пурных бактерий - всего лишь - 100 мВ!). При столь
большом отрица тельном потенциале становится возможным прямое использование
электронов от первичного акцептора для восстановления ферредоксина и
пиридиннуклеотида (рис. 12.17). Таким образом, восстановительную силу
Chlorobiaceae, возможно, получают не путем обратного транспор та электронов,
требующего затрат энергии. Такая независимость от обратного транспорта
электронов была бы важной отличительной чер той фотосинтеза у зеленых бактерий
по сравнению с пурпурными. Тог да фотореакция у Chlorobiaceae не уступала бы по
своей эффективности первой фотореакции цианобактерий. С эволюционной точки
зрения фо тосинтез зеленых бактерий мог бы быть связующим звеном между фо
тосинтезом пурпурных бактерий и фотосинтезом цианобактерий и растений. 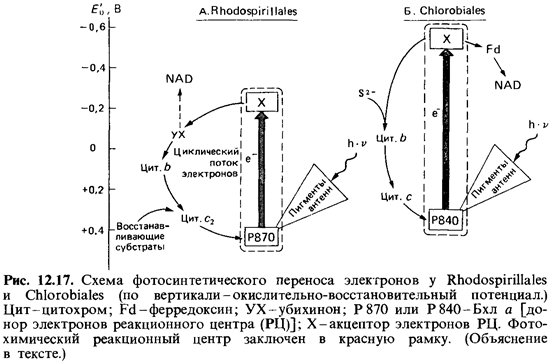
Заключение.
В процессе фотосинтеза происходит превращение энер гии света в
биохимическую энергию. Первичное действие света состоит в том, что в
фотохимических реакционных центрах электроны донора переносятся на акцептор в
термодинамически невыгодном направлении. По крайней мере часть электронов
возвращается по электрон-транс портной цепи к реакционным центрам. Благодаря
особому расположе нию компонентов электрон-транспортной системы в мембране это
со провождается направленным Переносом протонов и созданием протон ного
потенциала. Таким образом, аппарат фотосинтеза - это прежде всего протонный
насос, приводимый в действие светом. Протонный по тенциал обеспечивает
возможность преобразования энергии путем фосфорилирования.
Синтез АТР происходит с помощью тех же в своей ос нове механизмов, что и в
мембранах аэробных бактерий или митохондрий. Что же касается преобразования
энергии света в биохи мически полезную энергию (АТР), то здесь нет
принципиального разли чия между фототрофными бактериями и зелеными растениями.
У пур пурных бактерий роль фотосинтеза, по-видимому, исчерпывается этим
преобразованием. У цианобактерий и зеленых растений можно видеть дальнейший
этап эволюции фотосинтеза. У них благодаря последова тельному включению двух
фотореакций энергетический уровень элек тронов в ходе первой реакции удается
поднять настолько, что становит ся возможным восстановление ферредоксина и
NADP. Вторая фото реакция позволяет использовать в качестве источника
электронов воду. В результате такой комбинации наряду с запасанием энергии
происхо дит восстановление NADP и выделение 02.
Процесс
фотосинтеза представляет собой химическую реакцию, на иболее часто
совершающуюся на нашей планете. Ему мы обязаны как непрерывным синтезом нового
органического материала, так и существованием таких видов ископаемого топлива,
как уголь, нефть и при родный газ. Поэтому те огромные усилия, которые
потребовались для раскрытия тайны фотосинтеза, можно считать вполне
оправданными. Однако многие проблемы еще не решены. Поэтому наряду с экспери
ментально доказанными фактами в описанную выше модель фотосин теза был включен
ряд гипотез, для проверки которых нужны дальней шие настойчивые исследования. | 



