Описанные
выше бактериофаги, как правило, лизируют зараженные ими бактерии, и потому их
называют вирулентными. Некоторые фаги, однако, заражают
бактерий-хозяев, но не размножаются в них автономно и не вызывают лизиса. Такие
фаги называются умеренными. Видимо, их размножение происходит
синхронно с размножением бактерии. Лишь очень редко, в одной из 102-105 таких «лизогенных» бактерий,
фаг начинает спонтанно размножаться и клетка подвергается лизису. В этом случае
для того, чтобы обнаружить выход инфекционного фага, в качестве индикатора
нужен другой бактериальный штамм, для которого этот фаг вирулентен. Если смешать
лизогенные бактерии с избытком бактерий-индикаторов и посеять смесь на
агаризованную среду, то будут расти также и колонии лизогенных бактерий. Время
от времени некоторые клетки будут лизироваться и выходящие из них фаговые частицы,
будут заражать находящиеся по соседству чувствительные (индикаторные) бактерии.
Это приведет к появлению бляшек в сплошном бактериальном газоне. Однако в
середине каждой такой бляшки сохранится колония лизогенной бактерии (рис.
4.12). 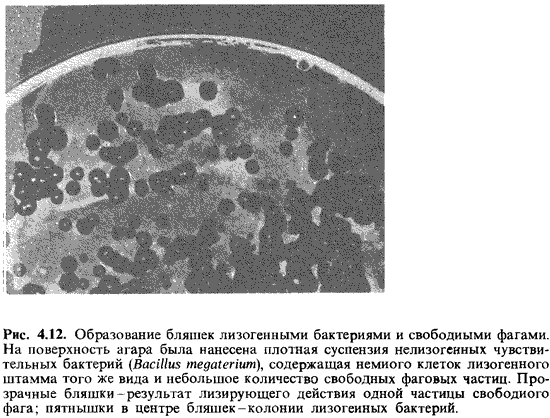
Лизогенные
бактерии обладают потенциальной способностью продуцировать фаги, но эту
способность нельзя обнаружить, ни морфологическим, ни серологическим
исследованием. Фаг в таком неинфекционном состоянии, передающийся только
дочерним клеткам при делении, называют профагом. Подобно
другим признакам бактериальной клетки, наличие в ней профага наследуется.
Поскольку все потомство лизогенной клетки тоже лизогенно, профаг, очевидно,
должен реплицироваться синхронно и регулярно вместе с хромосомой клетки-хозяина
(рис. 4.13). 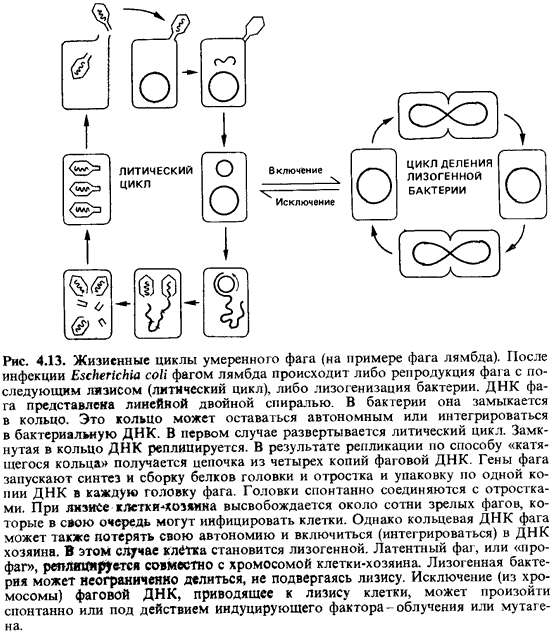
Лизогенные
бактерии иммунны к заражению теми фагами, которые присутствуют в них в виде
профага. Обеспечиваемый профагами иммунитет обусловлен не невозможностью
адсорбции (как при устойчивости к вирулентным фагам), а образованием особого
цитоплазматического белка-репрессора, препятствующего размножению вегетативных
фагов. Этот же репрессор препятствует обратному переходу профага в вегетативное
состояние и подавляет синтез фаговых белков. Возникновение лизогенного
состояния связано, таким образом, с образованием репрессорного белка.
Спонтанно,
без воздействия извне лизогенные бактерии лизируются редко. Однако целый ряд
факторов (ультрафиолетовые лучи, митомицин С или алкилирующие агенты) может
индуцировать в каждой клетке развитие профага, ведущее к образованию и
высвобождению инфекционного фага. Успех такой индукции зависит от
генетической конституции профага, физиологического состояния хозяина и условий
культивирования. Индукция связана, очевидно, с устранением или инактивацией
имеющихся молекул репрессора. Некоторые мутанты умеренных фагов образуют
термолабильный репрессор, и тогда достаточно уже повышения температуры до 44°С,
чтобы вызвать лизис бактерий.
Интеграция
и индукция фага λ (лямбда).
Изучение фага лямбда (λ), лизогенного
для Escherichia coli K12, позволило выяснить, каким образом
профаг связан с бактериальной хромосомой. Лизогенизация бактерий этим фагом
может служить примером жизненного цикла умеренного бактериофага. Длина
хромосомы фага лямбда оставляет всего 2% длины бактериальной хромосомы.
В свободных
фаговых частицах ДНК присутствует в виде линейной (не кольцевой) двойной
спирали (рис. 4.14). Каждая из цепей на одном конце выступает за пределы
дуплекса на 12 нуклеотидов. Эти два одноцепочечных конца комплементарны друг
другу; путем спаривания оснований они могут соединяться друг с другом, поэтому
их называют «липкими» концами. Если поместить такие молекулы ДНК in vitro в
раствор, то благодаря взаимодействию между комплементарными основаниями
одноцепочечных концов наступает равновесие между линейными и кольцевыми ДНК.
Такое же замыкание в кольцо происходит после того, как фаг лямбда инфицирует
клетку. При этом оба разрыва между концами цепей закрываются полинуклеотидлигазой.
Функция этого бактериального фермента состоит в том, чтобы устранять разрывы в
отдельных цепях двойных спиралей ДНК путем связывания нуклеотидов. Таким
образом, для замыкания линейной ДНК фага в кольцо никакие фаговые ферменты не
нужны. 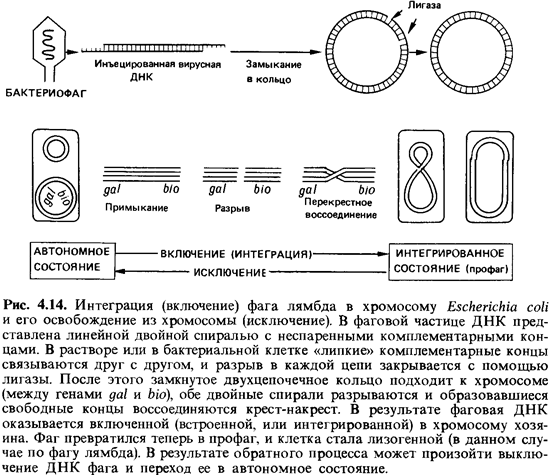
В лизогенных
клетках профаг прочно связан с хромосомой клетки-хозяина. При конъюгации клеток
профаг вместе с хромосомой хозяина переносится из клетки-донора в
клетку-реципиент. Генетические эксперименты показывают, что фаг лямбда
присоединен к хромосоме хозяина в совершенно определенном месте (между
галактозным опероном и биотиновым локусом). Вначале предполагали, что ДНК
бактериофага только прикрепляется к хромосоме бактерии в этом участке. Однако в
результате составления генетических карт фага, а также из опытов по
рекомбинации стало ясно, что фаговая ДНК при лизогенизации не просто
прикрепляется к бактериальной ДНК, а включается в нее.
Включение (интеграция)
ДНК профага в хромосому клетки-хозяина происходит, очевидно, в результате
разрыва и перекрестного воссоединения (рис. 4.14). За эту реакцию ответствен
фермент, названный лямбда-интегразой. Он узнает две разные, негомологичные
последовательности нуклеотидов - одну в хромосомной ДНК и одну в ДНК фага - и
тесно сближает обе двойные спирали друг с другом; затем последние разрываются и
снова соединяются крест-накрест. Отдельные этапы этой сайт-специфической
рекомбинации показаны на рис. 4.14.
В интегрированном
состоянии фаговая ДНК реплицируется вместе с бактериальной и подвержена тем же
регуляторным воздействиям, что и удвоение бактериальных хромосом. Информация,
содержащаяся в фаговой ДНК, в это время не проявляется. Только в результате
перехода профага в вегетативное состояние восстанавливается автономия фаговой
ДНК и начинается размножение фага. Этот обратный процесс может произойти
спонтанно или в результате индукции (например, под действием ультрафиолетового
облучения).Исключение фаговой ДНК из бактериальной хромосомы
происходит, вероятно, путем обращения процессов, приведших к ее включению, и
осуществляется очень точно: более 99% фаговых частиц, освобождающихся из
лизогенных клеток, идентичны с исходным (инфицирующим) фагом. Это означает, что
фаговая ДНК при ее выключении выщепляется точно в том же месте, где происходила
интеграция. Только в редких случаях (одном из 100 000) выключение ДНК фага
происходит аномально.
Как только
профаг в результате выключения перешел в вегетативное состояние, он опять
становится автономным и может размножаться в бактериальной клетке как
вирулентный фаг. Выключение, таким образом, приводит к лизису
бактерии и высвобождению фага лямбда. | 



