Репродукция
вируса в клетке-хозяине - процесс очень сложный. Его от дельные этапы, от
заражения клетки-хозяина до освобождения зрелых инфекционных частиц, довольно
хорошо изучены с биохимической, генетической и морфологической стороны на
примере фагов серии Т (Т2, Т4, Т6). Благоприятной предпосылкой для таких
исследований явилось то, что в фаговой ДНК вместо цитозина содержится
5-гидроксиметил-цитозин, и поэтому ее синтез легко проследить по появлению
этого основания. Кроме того, можно получить мутантные формы фага, у которых та
или иная стадия процесса репродукции блокирована или же протекает только в
определенных условиях. С помощью таких мутантов удалось выяснить, как
происходит внутри клетки-хозяина морфологическое развитие (морфопоэз) фага,
т.е. в какой временной последовательности синтезируются и соединяются различные
компоненты фаговых частиц. Подобно
другим вирусам, фаги неподвижны. При смешивании взвеси свободных фагов со
взвесью бактерий фаговые частицы в результате случайных столкновений с клетками
прикрепляются к поверхности последних (адсорбция) и вводят в клетку свою ДНК
(инъекция). По прошествии некоторого времени, необходимого для процессов
синтеза и созревания, клетки лизируются и новообразованные фаговые частицы
выходят наружу (см. рис. 4.13). 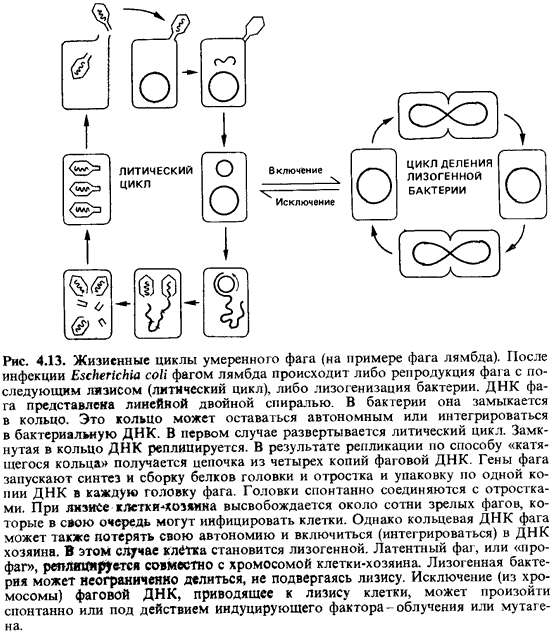
Адсорбция. Не
всякий фаг адсорбируется на любой бактерии. Специфичность отношений хозяина и
фага определяется специфичностью адсорбции, которая зависит от рецепторов,
имеющихся в клеточной стенке. Рецепторы для одних фагов находятся в
липопротеиновом слое, для других - в липополисахаридном. Фагорезистентность
некоторых бактерий определяется, вероятно, отсутствием у них соответствующих
рецепторов. При избытке бактериофага на одной клетке может адсорбироваться
200-300 фаговых частиц.
Внутриклеточное
развитие фага. За адсорбцией следует инъекция, т. е. введение ДНК в
клетку. У фага Т2 при этом базальная пластинка, по-видимому, фиксируется на
клетке, чехол отростка сокращается и в результате этого полый стержень входит
в бактериальную клетку. Опыты с фагом, у которого нуклеиновая кислота была
помечена 32Р, а белок - 35S, показали, что в клетку
проникает только нуклеиновая кислота, а белковая оболочка остается снаружи.
Можно было отделить эту оболочку от зараженной клетки без всякого ущерба для
размножения фага. Во время так называемого латентного периода, продолжающегося
у Escherichia coli в среднем 25 мин, в искусственно
разрушенных бактериальных клетках не удается обнаружить фага. Инъецированная
ДНК фага прежде всего вызывает полную перестройку метаболизма зараженной клетки
(рис. 4.10). Сразу же прекращается синтез бактериальной ДНК. Через несколько
минут прекращается также синтез бактериальной РНК и бактериальных белков, хотя
общее количество белка продолжает непрерывно возрастать. Синтез ДНК
возобновляется, даже с повышенной скоростью. Сначала фаговая ДНК образуется за
счет распавшейся бактериальной. Эту перестройку и последующее новообразование
фаговой ДНК можно количественно проследить по увеличению количества
5-гидроксиметилцитозина - основания, специфичного для ДНК некоторых Т-фагов.
Необходимые для синтеза фаговой ДНК ферменты образуются уже вскоре после
заражения; это так называемые «ранние белки». К «поздним белкам» относятся
белки оболочки и фаговые лизоцимы, или эндолизины; они образуются лишь во
второй половине скрытого периода. 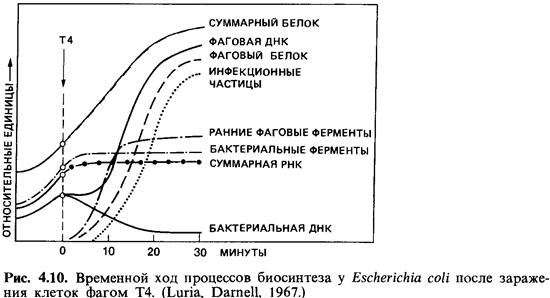
Заключительный
процесс – созревание - состоит в соединении фаговой ДНК с белком
оболочки и образовании зрелых инфекционных фаговых частиц. Созревание Т-фагов -
сложный многоступенчатый процесс. Сначала образуются капсиды, наполненные внутри
белками. После растворения этих внутренних белков готовые головки наполняются
ДНК до определенной, зависимой от типа фага плотности и закрываются. После
этого пристраиваются компоненты отростка. Последовательность этих процессов
можно проследить на условно летальных мутантах, у которых при 25°С все процессы
синтеза протекают нормально, а при 43°С тот или иной этап блокируется.
В конце
концов клеточная стенка бактерии размягчается под действием фагового лизоцима,
и новые фаги освобождаются. Такое внезапное разрушение клетки можно наблюдать
под микроскопом в условиях темного поля. Продолжительность латентного периода и
величина урожая фаговых частиц варьируют в широких пределах в зависимости от
вида фага, вида бактерии и условий среды (рис. 4.11). Удалось инфицировать
такие бактерии, как Haemophilus influenzae и Bacillus subtilis, нативной
ДНК, выделенной из бактериофага. Подобную инфекцию, соответствующую
генетической трансформации, называют трансфекцией. 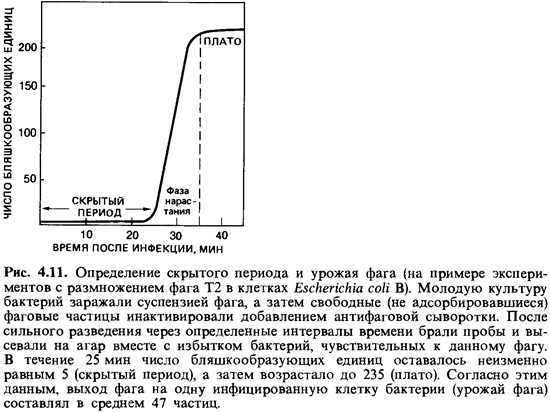
| 



