Распространение,
методы выделения и систематика. Пропионовокислые бактерии обитают в
рубце и кишечнике жвачных животных (коров, овец); они участвуют там в
образовании жирных кислот, главным образом пропионовой и уксусной. Благодаря им
образующаяся в рубце в результате различных видов брожения молочная кислота
превращается главным образом в пропионовую. Пропионовокислые бактерии не
встречаются в молоке, и их не удается выделить из почвы или при родных вод. Для
получения накопительной культуры этих бактерий питательную среду с лактатом и
дрожжевым экстрактом инокулируют швейцарским сыром и инкубируют в анаэробных
условиях. В швейцарский сыр, в созревании которого пропионовокислые бактерии
играют важную роль (определяя также его вкус), они попадают с сычужным
ферментом, применяемым в процессе приготовления сыра для свертывания молока.
Сычужный фермент добавляют в виде водного экстракта из телячьих желудков,
содержащего множество жизнеспособных пропионовокислых бактерий. Различают
несколько видов таких бактерий, из которых наиболее известны вид Propionibacterium freudenreichii и
его подвид shermanii, а также P. acidi-propionici(прежнее
название -Р. pentosaceum). Возбудитель угрей - воспаления
волосяных фолликулов у человека - тоже относится к пропионовокислым бактериям (P. acnes). Помимо
рода Propionibacterium к бактериям, образующим пропионовую
кислоту, причисляют Veillonella alcalescens (= Micrococcus lactilyticus),
Clostridium propionicum, Selenomonas и Micromonospora. Пропионат
как продукт брожения выделяют и многие другие бактерии.
Рост и
метаболизм представителей рода Propionibacterium. Этот род
относят к коринеформным бактериям; он включает грам-положительные
неподвижные палочки, не образующие спор. При неблагоприятных условиях роста
часто появляются булавовидные формы. Пропионовокислые бактерии на твердых
средах при доступе воздуха не растут. Так как они не переносят присутствия
атмосферного О2, способны расти в анаэробных условиях и
регенерировать АТР за счет энергии брожения, их считали организмами, облигатно
осуществляющими брожение. С другой стороны, давно известно, что они обладают
гемсодержащими ферментами, такими как цитохромы и каталаза. Действительно,
исследованные представители рода Propionibacterium оказались
способными расти как в анаэробных, так и в аэробных условиях. При достаточно
слабой контролируемой аэрации урожай их клеток во много раз больше, чем в
строго анаэробных условиях. В случае аэробного роста, правда, скорость диффузии
кислорода из газовой фазы в бактериальную взвесь не должна превышать его
потребления в процессе дыхания: кислород при измеримых парциальных давлениях
оказывает токсическое действие. Виды Propionibacterium, таким
образом, следует относить к микроаэротолерантным формам. В анаэробных условиях
представители этого рода сбраживают глюкозу, сахарозу, лактозу и пентозы, а
также лактат, малат, глицерол и другие субстраты с образованием пропионовой
кислоты. Расщепление гексоз идет по фруктозодифосфатному пути.
Исследуя
сбраживание глицерола клетками Propionibacterium acidi-propionici, Вуд
и Веркман (1936) нашли, что при этом происходит фиксация СО 2,
поддающаяся количественному определению. Связанный углерод можно было
обнаружить в выделяемом сукцинате, что указывало на карбоксилирование пирувата
с образованием дикарбоновых кислот. Такое карбоксилирование получило название
реакции Вуда- Веркмана. Эта реакция свойственна не только пропионовокислым бактериям
- она широко распространена у животных, растений и различных гетеротрофных
микроорганизмов. Соответствующие биохимические превращения описаны в разделе
7.5-там, где речь идет о глюконеогенезе.
Образование
пропионовой кислоты (метилмалонил-СоА-путь). Пропионовая кислота
образуется из молочной согласно следующему уравнению:
ЗСН3-СНОН-СООН
-*-* 2СН3-СН2-СООН + СН3-
СООН + С02 + Н20
Восстановление
лактата или пирувата до пропионата идет по пути, который по характерному
промежуточному продукту был назван метилмалонил-СоА-путем (рис. 8.3). Сначала
пируват при участии комплекса биотин-СО2
карбоксилируется метилмалонил-СоА-карбокситрансферазой с
образованием оксалоацетата, а затем восстанавливается через малат и фумарат до
сукцината. Транспорт электронов на этом этапе сопряжен с фосфорилированием.
Затем сукцинат с помощью СоА-трансферазы (сукцинил-СоА:
пропионат-СоА-трансферазы) присоединяется к СоА и таким образом активируется.
Сукцинил-СоА под действием метилмалонил-СоА-мутазы и при участии кофермента В12 (цианкобаламина)
превращается в метилмалонил-СоА, и только от этого промежуточного продукта,
наконец, отщепляется СО2; в результате образуется пропионил-СоА, а СО2связывается
с упомянутой выше метилмалонил-СоА-карбокситрансферазой. Из пропионил-СоА
образуется пропионат, в результате того что СоА-трансфераза переносит СоА на
сукцинат. Следует особо отметить, что в процессе образования пропионата две
группы (СО2 и СоА) пере носятся с последующего продукта на
предшествующий, не освобождаясь. Заслуживает внимания и участие трех кофакторов
(биотина, СоА и кофермента В12) в этом процессе. По метилмалонил-СоА
- пути пропионат образуется у большинства пропионовокислых бактерий, а также у Veillonella alcalescens и Selenomonas ruminantium. 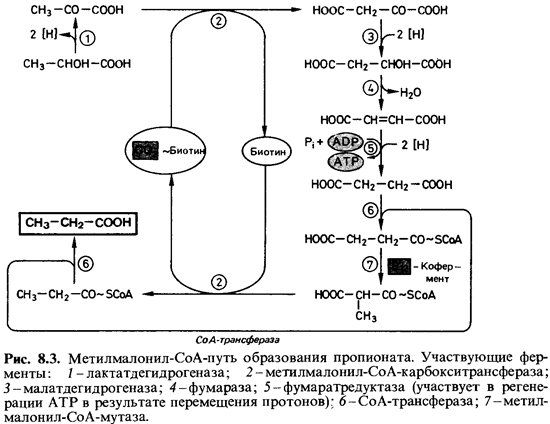
Реакции
метилмалонил-СоА-пути могут протекать и в обратном на правлении, как, например,
при расщеплении валина, изолеицина и жирных кислот с длинной цепью и нечетным
числом углеродных атомов. Из жирных кислот и изолеицина образуется
пропионил-СоА, который карбоксилируется с образованием метилмалонил-СоА. Для
пре вращения последнего в сукцинил-СоА при участии метилмалонил-СоА-мутазы
необходим витамин В12 (кофермент В12); это относится
и к корневым клубенькам бобовых, и к клеткам Rhizobium, и к
животным клеткам.
Акрилоил-СоА-путь. Образование
пропионовой кислоты у Clostridium propionicum, Bacteroides ruminicola и
Megasphaera elsdenii происходит более простым путем.
Промежуточным продуктом здесь является СоА-производное акриловой кислоты - акрилоил-СоА:
| 



