При внесении
бактерий в питательную среду они обычно растут до тех пор, пока содержание
какого-нибудь из необходимых им компонентов среды не достигнет минимума, после
чего рост прекращается. Если на протяжении этого времени не добавлять
питательных веществ и не удалять конечных продуктов обмена, то получим так
называемую периодическую культуру (популяцию клеток в ограниченном жизненном
пространстве). Рост в такой «закрытой системе» подчиняется закономерностям,
действительным не только для
одноклеточных, но и для многоклеточных
организмов. Периодическая культура ведет себя как многоклеточный организм с
генетически ограниченным ростом.
Кривая,
описывающая зависимость логарифма числа живых клеток от времени, называется кривой
роста. Типичная кривая роста (рис. 6.6) имеет S-образную форму и позволяет
различить несколько фаз роста, сменяющих друг друга в определенной
последовательности и в боль шей или меньшей степени выраженных: начальную (или
лаг-) фазу, экспоненциальную (или логарифмическую) фазу, стационарную фазу и фазу
отмирания. 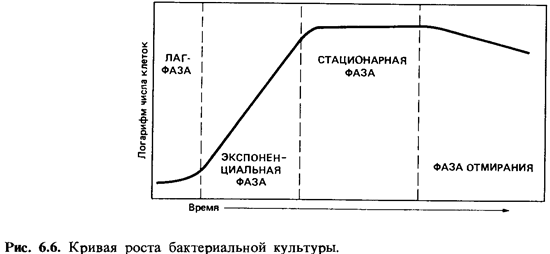
Рост микроорганизмов
на твердых питательных средах протекает в основном так же, хотя при этом
достигаются значительно более высокие плотности клеток.
Начальная
фаза. Эта фаза охватывает промежуток времени между инокуляцией и
достижением максимальной скорости деления. Продолжительность этой фазы зависит
главным образом от предшествовавших условий культивирования и возраста
инокулята, а также от того, насколько пригодна для роста данная среда. Если
инокулят взят из старой культуры (в стационарной фазе роста), то клеткам
приходится сначала адаптироваться к новым условиям путем синтеза РНК, образования
рибосом и синтеза ферментов. Если источники энергии и углерода в новой среде
отличаются от тех, какие были в предшествующей культуре, то приспособление
(адаптация) к новым условиям может быть связано с синтезом новых ферментов,
которые ранее не были нужны и поэтому не синтезировались. Образование новых
ферментов индуцируется новым субстратом. 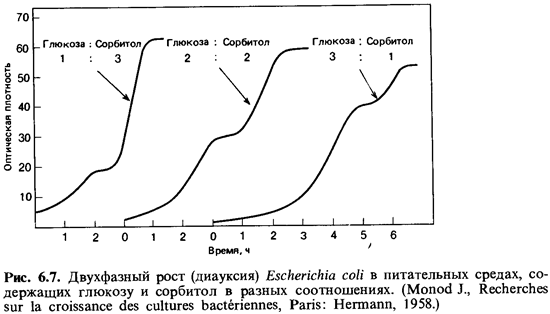
Хорошим
примером влияния субстрата на синтез ферментов служит так называемая диауксия (рис.
6.7). Это явление двухфазного роста или двойного цикла роста наблюдается на
средах, содержащих смесь питательных веществ. Из смеси глюкозы и сорбитола Escherichia coli, например,
поглощает в первую очередь глюкозу. Глюкоза индуцирует сначала в клетках синтез
ферментов, которые нужны для ее использования и одновременно подавляет
(репрессирует) синтез ферментов, необходимых для использования сорбитола. Эти
последние ферменты образуются лишь после того, как вся глюкоза будет
израсходована. Такие регуляторные процессы достаточно хорошо объясняют наличие
двух начальных фаз.
Количественное
изменение состава бактериальной клетки во время начальной фазы роста сильнее
всего затрагивает рибонуклеиновую кислоту: содержание РНК повышается в 8-12
раз. Это указывает на участие РНК и рибосом в синтезе ферментных белков.
Экспоненциальная
фаза. Экспоненциальная (логарифмическая) фаза роста характеризуется
постоянной максимальной скоростью деления клеток. Эта скорость во время
экспоненциальной фазы зависит от вида бактерий, а также от среды.
Энтеробактерии делятся через каждые 15-30 мин, Escherichia coli при
37°С - примерно каждые 20 мин. У других бактерий время генерации значительно
больше: у многих почвенных видов оно достигает 60-150 мин, а у Nitrosomonas и Nitrobacter
- даже 5-10 ч.
Величина
клеток и содержание в них белка у многих бактерий тоже остаются в
экспоненциальной фазе постоянными. В известном смысле можно сказать, что
бактериальная культура в этом случае состоит из «стандартных клеток». Если
точно установлено, что число клеток, содержание в них белка и их сухая биомасса
увеличиваются с одинаковой скоростью, то за ростом культуры можно следить,
пользуясь каким-нибудь одним из этих показателей.
Нередко,
однако, и в экспоненциальной фазе роста клетки периодической культуры
претерпевают изменения, так как постепенно изменяется среда: уменьшается
концентрация субстрата, увеличивается плотность клеточной суспензии и
накапливаются продукты обмена. В связи с тем, что в экспоненциальной фазе
скорость деления клеток относительно постоянна, эта фаза наиболее удобна для
определения скорости деления (и скорости роста). Изучая влияние факторов среды
(рН, окислительно-восстановительного потенциала, температуры, аэрации и т. д.),
а также пригодность различных субстратов, следят за увеличением числа клеток
или за мутностью (экстинкцией) клеточной суспензии во время экспоненциального
роста.
Стационарная
фаза. Стационарная фаза наступает тогда, когда число клеток перестает
увеличиваться. Скорость роста зависит от концентрации субстрата - при
уменьшении этой концентрации, еще до полного использования субстрата, она
начинает снижаться. Поэтому переход от экспоненциальной фазы к стационарной
происходит постепенно. Скорость роста может снижаться не только из-за нехватки
субстрата, но также из-за большой плотности бактериальной популяции, из-за
низкого парциального давления О2 или накопления токсичных
продуктов об мена; все эти факторы вызывают переход к стационарной фазе. И в
стационарной фазе могут еще происходить такие процессы, как использование
запасных веществ, распад части рибосом и синтез ферментов. Наблюдаемая картина
зависит от того, какой именно фактор лимитирует рост. Быстро гибнут лишь очень
чувствительные клетки; другие еще долго сохраняют жизнеспособность - до тех
пор, пока есть возможность получать необходимую для этого энергию в процессе
окисления каких-либо запасных веществ или клеточных белков.
Количество биомассы,
достигнутое в стационарной фазе, называют выходом или урожаем. Урожай зависит
от природы и количества используемых питательных веществ, а также от условий
культивирования.
Фаза
отмирания. Фаза отмирания и причины гибели бактериальных клеток в нормальных
питательных средах изучены недостаточно. Сравнительно легко понять случаи,
когда в среде накапливаются кислоты (при росте Escherichia, Lactobacillus). Число
живых клеток может снижаться экспоненциально. Иногда клетки лизируются под
действием собственных ферментов (автолиз). | 



