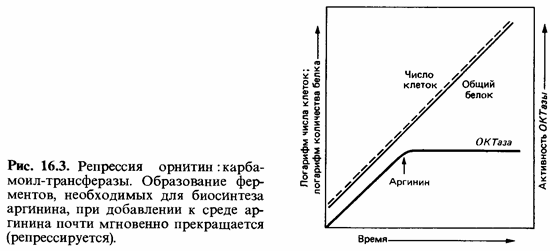
Репрессия конечным продуктом. Влияние конечного продукта биосинте тического пути на концентрацию ферментов этого пути можно проил люстрировать на примере синтеза аргинина (рис. 16.3). Аргинин синтезируется из глутамата (см. рис. 7.17) через орнитин, цитруллин и аргининосукцинат:
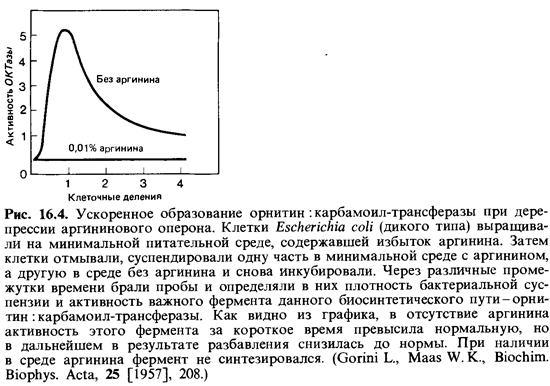
При выращивании клеток Escherichia coli на минимальной среде кон центрация одного из ферментов биосинтеза аргинина - орнитин :карба-моил-трансферазы (ОКТазы)-оказывается «обычной». При добавлении к минимальной среде аргинина (20 мкг/мл) образование ОКТазы сразу же прекращается вследствие репрессии. В результате дальнейшего роста клеток в присутствии аргинина происходит «разбавление» фермента, и его активность падает до очень низкого уровня. Если теперь отмыть клетки от аргинина и перенести их в минимальную среду, не содержа щую этой аминокислоты, то сразу же произойдет дерепрессия и синтез ОКТазы возобновится. Спустя короткое время концентрация фермента во много раз превысит его содержание в исходной суспензии клеток (рис. 16.4).
Лишь постепенно, после того как синтез анаболических ферментов приведет к образованию и накоплению аргинина, активность ОКТазы вновь снизится и достигнет «обычного» уровня.
На мутантах, дефектных по одному из ферментов биосинтеза аргинина, путем опытов в хемостате удалось показать, что состояние дерепрессии может сохраняться: если рост клеток лимитируется аргини ном, активность фермента в 25 раз выше «обычного» уровня.
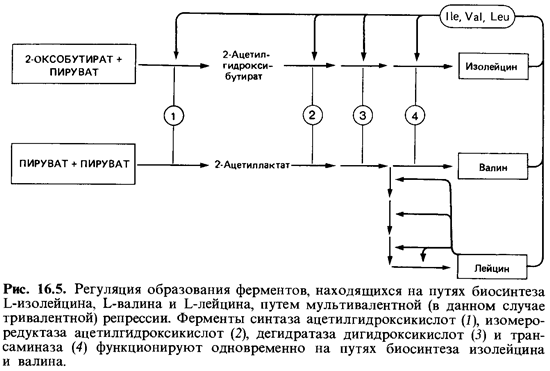
Схемы регуляции при разветвленных путях биосинтеза. Регуляция образования ферментов, участвующих в разветвленных путях биосинте за, очень сложна. Примерами могут служить системы, синтезирующие «семейство ароматических аминокислот», «семейство аспарагиновой кислоты» и «семейство пировиноградной кислоты» (см. рис. 7.17). Оче видно, что каждый конечный продукт может репрессировать образова ние ферментов только специфического пути биосинтеза. Ферменты, на ходящиеся перед местом разветвления путей, подвержены репрессии всеми конечными продуктами, действующими одновременно (мультива-лентная репрессия). Синтез этих ферментов подавляется лишь тогда, когда в питательной среде присутствуют все конечные продукты; если же добавлять их по отдельности, они такого эффекта не оказывают.
Интересный пример разветвленного пути биосинтеза-образование L-изолейцина, L-лейцина и L-валина. В роли катализатора четырех эта пов биосинтеза валина из пирувата и изолейцина из 2-оксобутирата вы ступает на каждом этапе в обеих цепях один и тот же фермент (рис. 16.5), а образующийся 2-оксоизовалерат (2-оксо-З-метилбутират) служит общим предшественником валина и лейцина.
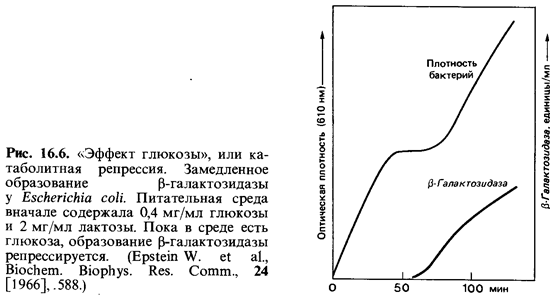
Катаболитная репрессия. В то время как репрессия конечным продук том действует на путях биосинтеза, при помощи катаболитной репрес сии регулируются катаболические реакции. Если в питательной среде содержатся два разных субстрата, то, как правило, бактерии «предпочи тают» тот, который обеспечивает более быстрый рост. Этот субстрат вызывает репрессию синтеза тех ферментов, которые нужны для ис-
пользования второго субстрата (рис. 16.6). Катаболитная репрессия ле жит в основе, например, известного явления диауксии (см. рис. 6.7). При наличии двух субстратов (глюкоза + сорбит, глюкоза + ацетат) они используются клетками Е. coli не одновременно, а последовательно. Сна чала используется глюкоза, и при этом она подавляет синтез фермен тов, необходимых для расщепления второго субстрата.
Сложнее обстоит дело, когда та или иная аминокислота служит для клеток не только источником энергии и углерода, но также источником
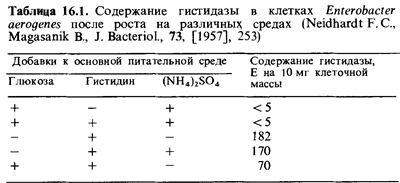
азота. На примере использования гистидина клетками Enterobacter aerogenes видно, как может влиять глюкоза на содержание ферментов, с помощью которых происходит усвоение какой-либо аминокислоты. Измеряли содержание в клетках гистидазы, взятой в качестве типичного катаболического фермента (табл. 16.1). В среде, содержавшей глюкозу и аммонийный азот, образования гистидазы не происходило даже в присутствии гистидина. Глюкоза и ионы аммония совместно почти полностью подавляли синтез этого фермента. При отсутствии ионов NH4, т.е. в условиях, когда клетки не имели дополнительного источ ника азота и должны были использовать азот гистидина, репрессирую щее действие глюкозы на синтез гистидазы было менее выражено. Этот пример показывает, что недостаток азота может частично снимать ка-таболитную репрессию, вызываемую глюкозой. Таким образом, синтез катаболического фермента зависит здесь не только от наличия индукто ра и источника энергии, но и от присутствия азота в среде.
Регулирование концентрации ферментов центральных метаболических путей. Эти ферменты тоже могут быть подвержены регуляции. В клет ках Escherichia coli, растущих в аэробных условиях, ферменты цикла трикарбоновых кислот присутствуют в высокой концентрации; между тем при росте в анаэробных условиях их активность в 10-20 раз мень ше, а образование 2-оксоглутаратдегидрогеназы бывает полностью по давлено. Некоторые анаплеротические ферменты, такие как малатсинта-за, изоцитрат-лиаза и глиоксилат-карболигаза, содержатся только в тех клетках, которым они необходимы для использования имеющегося субстрата.




