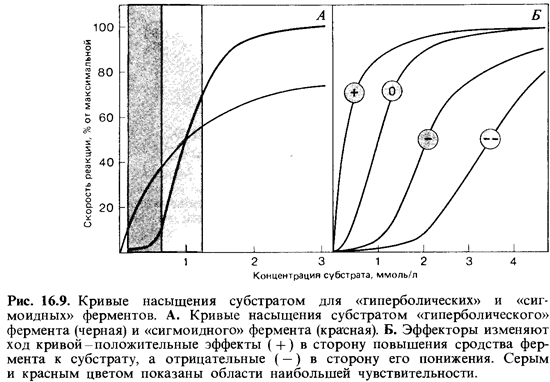
Простые ферменты. Для большинства ферментов характерна гипер болическая кривая насыщения субстратом. Скорость реакции зависит только от концентраций субстрата и продукта и гиперболически возра стает с повышением концентрации субстрата, т.е. удовлетворяет усло виям соотношения Михаэлиса-Ментен. Такие ферменты называют про стыми или «гиперболическими» ферментами (рис. 16.9, А, черная кри вая).
Понятно, что при высокой концентрации субстрата фермент перера батывает его быстрее, чем при более низкой концентрации. Насколько чувствительно реагирует фермент на изменение концентрации субстра та, зависит от крутизны кривой его насыщения субстратом. Чем круче кривая, тем больше повышается скорость реакции при незначительном сдвиге концентрации субстрата. Как видно из рис. 16.9, А, крутизна кри вой и соответственно чувствительность больше всего при низких кон центрациях субстрата. Из этих рассуждений ясно, что скорость оборота
веществ в клетке зависит от их концентрации. Как правило, субстраты ферментов (метаболиты) содержатся в клетке в концентрациях ниже Кт.
Регуляторные ферменты. Свойства регуляторных ферментов намного более сложны. Кривые насыщения субстратом для большинства этих ферментов отклоняются от гиперболической формы и часто становятся сигмоидными (рис. 16.9, А и J5). У таких кривых имеется область значи тельно большей крутизны, чем у кривых насыщения для простых фер ментов. В этой области, примерно между 1/2 и 1 */4 Кт, регуляторные ферменты очень чувствительны-даже небольшого изменения концен трации субстрата достаточно, чтобы сильно изменить скорость реакции.
Сигмоидная форма кривой указывает на то, что фермент построен из субъединиц, между которыми существуют кооперативные взаимодей ствия. Очевидно, связывание субстрата с каталитическим центром одной из субъединиц фермента повышает сродство к субстрату других участков связывания в той же молекуле. Регуляторные ферменты со стоят из двух или более, чаще всего из четырех, субъединиц.
Кроме каталитических центров, распознающих и связывающих суб страты, у регуляторных ферментов есть и другие стереоспецифические участки-так называемые аллостернческие центры. Это места связыванияэффекторов, изменяющих сродство фермента к субстрату. Имеются особые участки для связывания положительных эффекторов (активато ров) и для отрицательных эффекторов (ингибиторов). Под влиянием эф фекторов изменяется форма кривой насыщения (степень ее «сигмоидности» (рис. 16.9, Б). Говорят не только об аллостерических центрах, но
также об аллостерическом торможении и аллостерических ферментах
(термины «аллостерические ферменты», «регуляторные ферменты» и «сигмоидные ферменты» часто употребляют как синонимы).
Степень кооперативности выражают с помощью коэффициента кооператив-ности (или коэффициента Хилла)п. Он соответствует числу зависимых друг от друга областей связывания или числу субъединиц, участвующих в кооператив ной реакции. При отсутствии кооперативности п = 1. Для аллостерического фер мента, состоящего из четырех субъединиц, возможны значения и > 1 и п < 4 в за висимости от степени положительной кооперативности. При отрицательной кооперативности п < 1. Кроме того, имеются более сложные системы, на ко торых мы здесь останавливаться не будем.
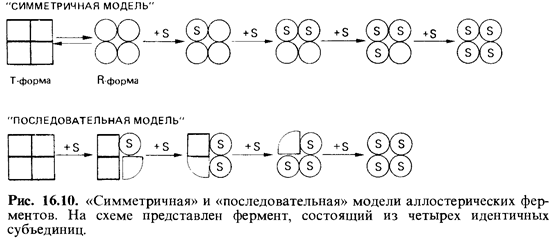
Модели кооперативности. Попытки объяснить кооперативность ме жду субъединицами ферментов, проявляющуюся в сигмоидной форме кривой насыщения субстратом, сводятся к двум гипотезам: «симметричной» модели (Моно и сотр.) и «последовательной» модели (Кош-ленд); они схематически представлены на рис. 16.10.
В основу обеих моделей положено представление о том, что фер менты могут существовать в различных формах-в активной форме (с высоким сродством к субстрату) и в неактивной (с малым сродством к субстрату). В каком соотношении между собой будут находиться раз ные формы фермента, зависит от наличия и концентрации лигандов (молекул субстрата, активаторов и ингибиторов). Разница между гипотезами касается того, как происходит конформационное изменение.
Согласно симметричной модели, фермент представлен только двумя конформационными состояниями, находящимися в динамическом рав новесии. При этом все субъединицы данной молекулы фермента находятся в одной и той же конформации; промежуточных состояний нет, существуют только симметричные олигомеры (рис. 16.11). Равновесие характеризуется аллостерической постоянной L. В отсутствие лигандов, как правило, неактивное основное состояние Т (от англ. tense-напря женный) преобладает над активным состоянием R (от англ. relaxed-расслабленный). При добавлении лигандов они реагируют с теми моле-
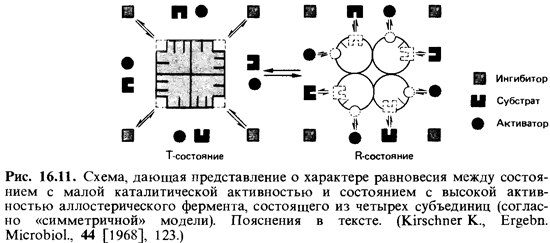
кулами фермента, которые находятся в соответствующей конформации: ингибиторы-с молекулами в состоянии Т, субстраты и активаторы-с молекулами в состоянии R. При связывании субстрата фермент удержи вается в состоянии R. Чтобы восстановить равновесие между двумя конформационными состояниями, часть других молекул фермента тоже переходит в состояние R. Поскольку каждая молекула имеет несколько участков для связывания субстрата, небольшого числа молекул субстра та достаточно для того, чтобы привести намного большее число таких участков в состояние высокой каталитической активности; таким обра зом, облегчается связывание и других молекул субстрата. В этом и со стоит кооперативный эффект, определяющий сигмоидную форму кри вой «концентрация субстрата-скорость реакции». Отрицательный эф фектор оказывает обратное действие.
В последовательной модели предполагается, что фермент приобре тает каталитически активную конформацию только в результате взаи модействия с субстратом (рис. 16.10). Если фермент состоит из несколь ких субъединиц, то конформационное изменение одной из них, вызванное субстратом, последовательно передается другим субъедини цам и облегчает им связывание добавочных молекул субстрата. Воз можно образование несимметричных олигомеров (на рис. 16.10 это те-трамеры) с субъединицами, имеющими разную конформацию. Присут ствие активаторов способствует переходу в активную форму, а от рицательные эффекторы его затрудняют.
Аллостерические ферменты и эффекторы. Регуляторные ферменты, как правило, имеются в каждом пути биосинтеза и в некоторых путях катаболизма. В большинстве случаев они находятся в начале цепи биосинтеза и занимают, таким образом, ключевую позицию.
Аллостерические эффекторы представляют собой низкомолеку лярные соединения - это либо конечные продукты биосинтеза, либо ве щества, концентрация которых может отражать состояние клеточного метаболизма, например ATP, ADP, AMP, ацетил-СоА, фосфоенолпиру-ват и NADH-,.
Изменение активности ферментов путем ковалентной модификации.
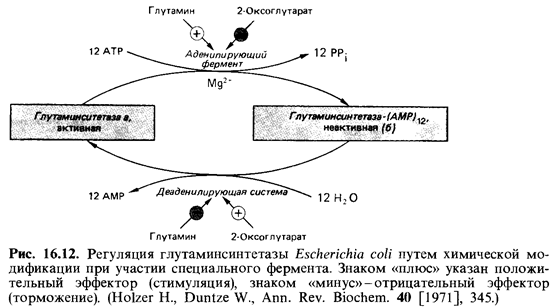
Для небольшого числа ферментов известен регуляторныи механизм иного типа: они могут изменяться под действием других ферментов. Это изменение может приводить к повышению или снижению активно сти фермента. Оно может состоять в аденилировании, фосфорилирова-нии или ацетилировании. Речь идет об относительно быстром процессе. У млекопитающих под воздействием ферментов активируются и инак-тивируются гликогенфосфорилаза и гликогенсинтетаза. У Escherichia coli не только подавляется образование глутаминсинтетазы, но и актив ность имеющегося фермента за несколько минут снижается на 80-90%, если в питательную среду добавить ионы аммония. Это снижение ак тивности обусловлено катализируемой особым ферментом химической модификацией: активная глутаминсинтетаза а в результате ее аденили-рования превращается в неактивную глутаминсинтетазу Ь (рис. 16.12). Наличие глутамина в клетке стимулирует аденилирующий фермент, а свободный 2-оксоглутарат оказывает противоположное действие. Пос ле удаления из среды ионов аммония в клетках создается недостаток глутамина, и глутаминсинтетаза вновь реактивируется в результате от щепления групп адениловой кислоты (AMP) под действием деаденили-рующей системы.




