Теоретически регуляция синтеза ферментов могла бы осуществляться как при транскрипции, так и при трансляции. Как выяснилось, экспрес сия генов у прокариот регулируется главным образом на уровне транскрипции. Большинство генов, кодирующих структуру полипептидных цепей, подвержено регуляции-точнее, регулируется их транскрипция. От условий среды и состояния клеточного метаболизма зависит, будут ли синтезироваться определенные ферменты, т.е. будут ли вообще и с какой частотой транскрибироваться соответствующие гены.
Для такой регуляции нужно, чтобы к ДНК из других частей клетки поступали определенные сигналы. Сигнальные вещества, или молекулы-эффекторы, представляют собой низкомолекулярные соединения, такие как сахара или их производные, аминокислоты или нуклеотиды. По скольку такие эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для каждого из них служит определенный регуля-торный белок. Если эффектор присутствует в клетке в высокой концен трации, то в результате специфического присоединения к регуляторному белку он изменяет его кон формацию и тем самым-его способность связываться с ДНК. Регуляторный белок, который связывается с ДНК в отсутствиеэффектора (индуктора), называют репрессором. Если же бе лок связывается с ДНК в присутствииэффектора (корепрессора), его на зывают апорепрессором.
Участки ДНК, к которым присоединяются регуляторные белки,-это не сами структурные гены, а непосредственно прилегающие к ним обла сти, называемые промоторами и операторами. Промотор представляет собой последовательность оснований, распознаваемую ДНК-зависимой РНК-полимеразой; он служит местом связывания РНК-полимеразы, и от него начинается транскрипция. С промотором связаны и гены, экспрессия которых не подвержена регуляции. Промоторы регулируемых генов могут изменять свои свойства в результа те связывания регуляторных белков. Оператор представляет собой нуклеотидную последовательность, расположенную между промото ром и структурными генами. Он тоже взаимодействует с регуля-торным белком-репрессором, от которого зависит, будет ли подавлена транскрипция или она произойдет. Промотор, оператор и структурные гены образуют оперон. Опероном называют группу функционально свя занных между собой генов. Белки, кодируемые генами одного оперона,-это, как правило, ферменты, катализирующие разные этапы одного метаболического пути. Транскрипция генов оперона ведет к синтезу одной общей (полицистронной) молекулы мРНК.
За синтез регуляторных белков ответственны гены-регуляторы, ко торые, вероятно, являются конститутивными. Они могут располагаться по соседству с соответствующим опероном, но это не обязательно.
По-видимому, РНК-полимераза способна правильно присоединяться к промотору только в форме полного голофермента. Она состоит из субъединиц а, р, р1, а и со. В отсутствие легко отделяющегося фактора сигма (а) фермент обладает полной каталитической активностью, но не способен связываться со специфическим участком ДНК - промотором. Этот фактор транскрипции (сигма) играет, вероятно, важную роль при специфическом присоединении полимеразы к ДНК.
За прекращение (терминацию) синтеза мРНК у конца оперона или отдельного гена, по-видимому, также ответственна специфическая область ДНК - терминатор. Какую роль играет фактор терминации (р)-тетрамерный белок-при отделении РНК-полимеразы от ДНК, по ка неясно.
Матричная РНК (мРНК) в отличие от тРНК и рРНК неустойчива и недолговечна; время ее полураспада составляет от 0,5 до 5 мин. Кон центрация той или иной мРНК в клетке зависит только от частоты транскрипции соответствующего гена; и в свою очередь она определяет содержание в клетке ферментов, кодируемых этим геном.
Различают индуцибельные и репрессибельные опероны. Опероны, управляющие катаболизмом лактозы, галактозы и арабинозы, являются индуцибельными, т. е. максимальная частота их транскрипции достигает ся только тогда, когда в питательной среде присутствует внешний эф фектор-лактоза, галактоза или арабиноза. Внешние эффекторы назы вают также внешними индукторами. Синтез ферментов индуцибельных оперонов включается посредством индукции. Наоборот, опероны, управ ляющие синтезомаргинина, гистидина или триптофана, являются ре-прессибельными, т.е. максимальная частота транскрипции достигается только при отсутствии в клетке соответствующих низкомолекулярных эффекторов-аргинина, гистидина и триптофана (или в том случае, если их концентрация ниже критического порогового уровня).
Такие эффекторы называют корепрессорами, а соответствующие ре-гуляторные белки - белками-репрессорами или апорепрессорами. Синтез ферментов репрессибельного оперона включается посредством дере-прессии.
Индукция лактозного оперона (отрицательный контроль). Лактозный оперон (/ас-оперон) Escherichiacoli содержит /ас-промотор, /ас-оператор и структурные гены для трех ферментов: $-галактозидазы, пермеазы и трансацетилазы (рис. 16.7). Этот оперон был тщательно исследован; удалось выделить его ДНК; была определена нуклеотидная последова тельность области промотор-оператор; были выделены и исследованы регуляторные белки.
Оперон находится под отрицательным контролем, т.е. регуляторный белок (/ас-репрессор) остается связанным с оператором и препятствует транскрипции до тех пор, пока отсутствует индуктор. Внешним индук тором служит лактоза (а-В-галактозил-р-1,4-В-глюкоза). Она транспор тируется с помощью пермеазы и превращается в аллолактозу (a-D-ra-лактозил-р-1,6-В-глюкозу), действующую как индуктор (внутренний). Катализатором этого превращения служит (3-галактозидаза. Оба фер мента -пермеаза и (3-галактозидаза-присутствуют и в неиндуциро-ванных клетках, но в концентрациях, составляющих менее 0,001 от их концентраций после полной индукции. Связав аллолактозу, /ас-репрес сор претерпевает конформационное изменение, уменьшающее его срод ство к ДНК оператора, и в результате освобождает оперон для транскрипции.
Лактозный оперон подвержен также второму контролю - положительному. Это означает, что транскрипция происходит лишь тогда, ког да с промотором связан второй регуляторный белок-САР, или CRP.
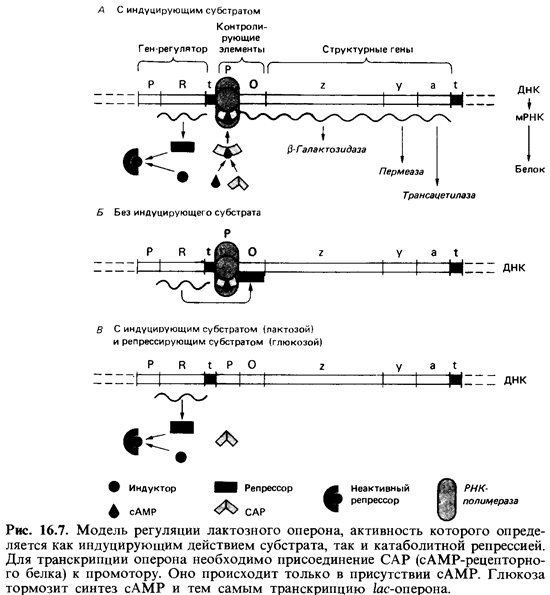
Сокращения CAP (catabolite activator protein) и CRP (cyclic AMP receptor protein) - синонимы. Связывание CAP служит необходимым ус ловием для присоединения РНК-полимеразы к ДНК. Однако САР мо жет связаться с промотором только в том случае, если в клетке в доста точно высокой концентрации присутствует циклический AMP (сАМР). Катаболитная репрессия /ас-оперона. Если в питательной среде для Escherichia coli содержатся лактоза и глюкоза, синтез ферментов /ас-опе рона подавляется (см. рис. 16.6 и 16.7). Такое действие глюкозы обусловлено тем, что в ее присутствии внутриклеточная концентрация сАМР остается низкой. Глюкоза (так же как фруктоза и глкжозо-6-фос-фат) репрессирует и другие индуцибельные пути катаболизма (расще пление арабинозы, галактозы, сорбитола, глицерола и др.).
Снижение концентрации сАМР под действием глюкозы обусловлено, вероятно, локализацией ферментааденилатциклазы, при участии кото рого образуется сАМР:
Фермент в клетке связан с мембраной. Его активность высока, если си стема транспорта Сахаров находится в фосфорилированной форме. Если же эта система затрачивает много энергии на транспорт Сахаров (что связано с их фосфорилированием), активность аденилатциклазы сни жается.
Индукция арабинозного оперона (положительный контроль). Араби-нозный оперон Е. coli подвержен похожительной регуляции, так же как рамнозный и мальтозный опероны. Он содержит структурные геныara В, ara А и ara D для ферментов, участвующих в превращении L-араби-нозы в D-ксилулозо-З-фосфат. Экспрессия оперона индуцируется араби-нозой. Как и многие другие системы катаболизма, оперон подвергается регуляции в области промотора, с которым связан САР, активиро ванный циклическим AMP. Кроме того, в этом опероне есть еще два ре гулируемых участка - оператор и инициатор. С оператором связан регу-ляторный белок, кодируемый геном ara С. Этот белок действует как репрессор-присоединяясь к оператору, он препятствует транскрипции. Однако в присутствии арабинозы он становится активатором-присое диняется к промотору и делает возможной транскрипцию. Таким обра зом, арабинозный оперон находится под влиянием как отрицательного, так и положительного контроля со стороны специфического белка-регу лятора.
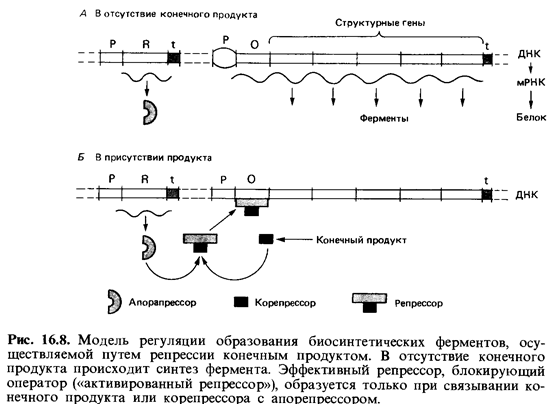
Репрессия триптофанового оперона конечным продуктом. Триптофа-новый оперон Е. coli содержит структурные гены для синтеза пяти фер ментов, участвующих в превращении хоризмата в триптофан, а также оператор и промотор в начале оперона. Второй, малоэффективный про мотор внутри серии структурных генов можно оставить без внимания. Детально изученная функция данного оперона полностью соответствует обобщенной модели, предложенной в 1961 г. Жакобом и Моно для ре-прессибельного оперона (рис. 16.8). Расположенный далеко от оперона ген-регулятор trp R кодирует эффекторный белок - апорепрессор. В при сутствии корепрессора - триптофана, имеющего высокое сродство к бел-ку-апорепрессору, репрессор подавляет транскрипцию оперона. Сниже ние концентрации триптофана вновь приводит к освобождению оператора и синтезу мРНК.
Аутогенная регуляция. Это такой механизм, в котором функцию бел ка-регулятора выполняет продукт одного из генов, находящихся в самом опероне. Принцип аутогенной регуляции состоит в том, что ре-гуляторный белок управляет транскрипцией оперона и тем самым влияет на свой собственный синтез. При этом безразлично, какому кон тролю подвергается оперон-положительному или отрицательному.
Наиболее изученной системой такого типа является регуляция ис-
пользования гистидина у Salmonella. Превращение гистидина в глута-мат, ионы аммония и формамид катализируют четыре фермента, структурные гены которых (гены hut) расположены рядом, образуя один оперон. Их экспрессия контролируется репрессорным белком. Ген, коди рующий этот белок, находится между структурными генами, т.е. в самом опероне. Этот оперон -hut-индуцируется первым продуктом катаболизма-уроканатом (индукция продуктом). В связи с этим индук ция оперона ведет не только к синтезу ферментов, расщепляющих ги-стидин, но и к накоплению белка-репрессора. Последний вновь притормаживает транскрипцию генов hut, включая ген-регулятор. Аутогенная регуляция создает возможность мягкого, плавного управления синтезом ферментов. Такого рода саморегулирующиеся системы широко распро странены у бактерий, эукариот и бактериофагов.
Регуляция синтеза рРНК и тРНК. В то время как синтез мРНК регу лируется путем индукции и репрессии (в частности, катаболитной), регу ляция синтеза стабильных РНК происходит совершенно иначе. Как уже давно известно, штаммы дикого типа Escherichia coli, Salmonella typhi-murium и Bacillus subtilis очень резко реагирует на недостаток какой-ли бо аминокислоты. Они сразу же перестают синтезировать не только белки, но и РНК. Эта строгая зависимость от наличия «строительных блоков» для биосинтеза белка-аминокислот-позволяет заключить, что синтез стабильных РНК зависит не только от наличия их собственных предшественников - нуклеотидов. Выделение мутантов с менее строгой, «расслабленной» регуляцией, синтезирующих РНК даже при отсутствии одной аминокислоты, привело к выводу, что в регуляции участвует по меньшей мере один ген (rel А). Если отсутствие какой-либо аминокис лоты в клетках дикого типа приводит к накоплению необычных нуклео-тидов (ppGpp и pppGpp), то у мутантов этого не наблюдается. Можно предположить, что эти нуклеотиды служат сигнальными веществами, приостанавливающими синтез рРНК и тРНК.




