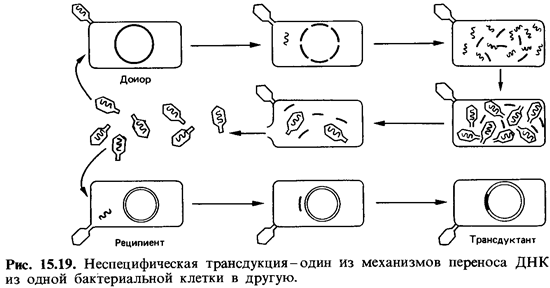
Неспецифическая трансдукция. Перенос участков бактериальной хро мосомы фагами был открыт в 1951 г. Ледербергом и Циндером у Salmonella typhimurium. В решающем эксперименте (рис. 15.19) штамм-донор В+инфицировали умеренным бактериофагом Р22. После лизиса клетки-хозяина выделяли свободные фаги и инкубировали их вместе со штаммом-реципиентом В ~, который генетически отличался от штамма В+ по меньшей мере одним признаком. Авторы нашли, что после вы сева инкубированных клеток на подходящую среду появлялись реком-бинанты, обладавшие признаками штамма-донора В + .
Процессы, происходящие при таком неспецифическом переносе ДНК, весьма сложны. Во время репродукции фага Р22 в клетках штам ма-донора В + в капсиды вместо фаговой ДНК могут включаться фраг менты бактериальной хромосомы. Таким образом, фаголизат содержит смесь нормальных и дефектных фагов. Заражение штамма-реципиента В" нормальным фагом ведет, как правило, к лизису клеток. Однако в некоторые клетки проникают дефектные трансдуцирующие фаги, ДНК которых способна рекомбинироваться с хромосомой реципиента. Происходит обмен гомологичными участками ДНК, что может приве сти к замене дефектного гена реципиента интактным геном донора.
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероят ность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 ~б до 10~8. Становится понятно, что с помощью одной частицы фага Р22 Salmonella или неспецифически трансдуцирующего фага PI Escherichiacoli в каждом случае может быть трансдуцирован только один ген (или несколько очень близко расположенных генов). Количество бактериальной ДНК, сравнимое с геномом фага, составляет лишь 1-2% всего количества ДНК, содержащегося в бактериальной клетке. Исключение составляет бактериофаг PBS 1Bacillus subtilis, который может трансдуцировать до 8% генома хозяина.
Специфическая трансдукция. Наиболее известным примером служит трансдукция, осуществляемая фагом X(см. разд. 4.2.2). Обычно он трансдуцирует лишь определенные гены, а именно gal и Ыо. Как уже говорилось, этот фаг при переходе в состояние профага включается в определенный участок хромосомы бактерии-хозяина - между генами gal и Ыо. Отделение фаговой ДНК от бактериальной хромосомы (например, в результате УФ-облучения) может произойти неточно, т.е. ка кой-то фрагмент ее останется в хромосоме, а близко расположенные гены клетки-хозяина будут захвачены фаговой ДНК. По-видимому, при чиной этого может быть неправильная рекомбинация.
В случае заражения трансдуцирующим фагом клеток, дефектных по определенному гену, например gal~, может произойти рекомбинация с заменой собственного дефектного гена бактерии интактным трансду-цированным геном; при этом образуются рекомбинанты (трансдуктанты) gal+.
Подобным же образом происходит перенос генов бактериофагом Phi 80. Его ДНК включается в хромосому вблизи генов, кодирующих ферменты, ответственные за биосинтез триптофана. По этой причине Phi 80 особенно пригоден для переноса генов trp.
Предпосылкой успешного переноса генов при специфической транс-дукции (в отличие от неспецифической) является интеграция фага в ге ном клетки-хозяина.
В некоторых случаях было показано, что трансдуцированный фрагмент ДНК не вступает в рекомбинацию с хромосомой реципиента, а остается вне хромосомы. В этом случае клетка становится гетерозиготной по перенесенным генам. Перенесенная ДНК транскрибируется (на это указывает синтез соответ ствующего генного продукта), но не реплицируется. Это приводит к тому, что при клеточном делении донорский фрагмент переходит только в одну из дочер них клеток (абортивная трансдукция). Если реципиент ауксотрофный, а перене сенный фрагмент исправляет соответствующий дефект, то расти могут только те клетки, которые унаследовали этот фрагмент; при посеве на агар они обра зуют мельчайшие колонии.