У эукариотических организмов в процессе оплодотворения объединяют ся гаплоидные наборы генов и образуется диплоидная зигота. В дочер нем диплоидном организме после немногих или же многократных ми-тотических делений в процессе редукционного деления (мейоза) происходит перекомбинирование хромосом, принадлежавших двум ро дительским наборам, и снова образуются клетки с гаплоидными наборами генов (гаметы). Такому половому способу «перетасовки» генетиче ского материала противопоставляют парасексуальные процессы, к которым относится и рекомбинация признаков у прокариот. Бактерии почти всегда гаплоидны; у них имеется только один набор генов. Зи готы образуются и у бактерий, но они никогда не бывают продуктом объединения целых клеток. Как правило, из клетки-донора в клетку-ре ципиент переносится лишь часть генетического материала, т.е. обра зуется неполная зигота (мерозигота). Хромосома реципиента спаривает ся с фрагментом хромосомы донора, и они обмениваются отдельными участками. При последующем делении ядра и клетки возникает клетка, содержащая только рекомбинированную хромосому (рис. 15.11). У бактерий известны три способа передачи признаков: конъюгация, трансдук-ция и трансформация.В результате этих трех процессов ДНК переносится из бактерии-до нора в бактерию-реципиент. Эти процессы отличаются друг от друга способом транспортировки ДНК. После переноса ДНК в клетке-реци пиенте происходит рекомбинация. При этом ДНК донора встраивается в ДНК бактерии-реципиента. Клетку, в которой произошла рекомбина ция, называют рекомбинантом. 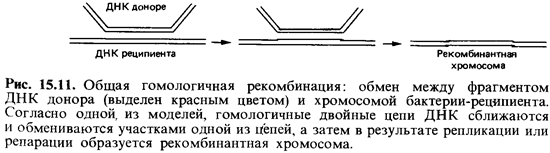
| 



