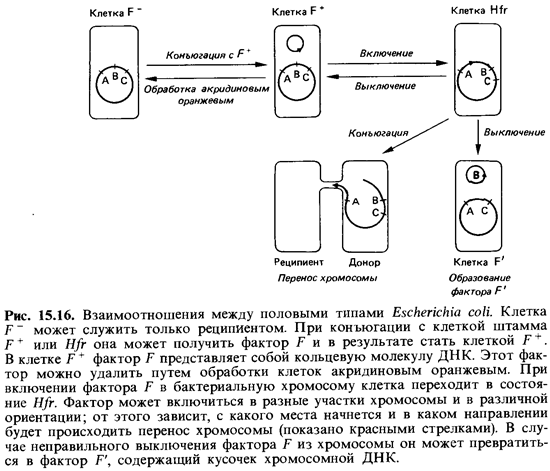
Фактор F и состояние Hfr. При исследовании процесса скрещивания бактерий выяснилось, что способность клетки быть донором связана с наличием особого фактора, который при конъюгации передается из одной клетки в другую - полового фактора F (от fertility - плодовитость). Клетки, не содержащие фактора F(клетки F~), могут функционировать только как реципиенты. При конъюгации, т.е. при прямом контакте между клетками, частота передачи фактора F близка к 100%. Таким образом, клетки-реципиенты в результате конъюгации превращаются в потенциальных доноров; при этом хромосомные признаки еще не передаются.
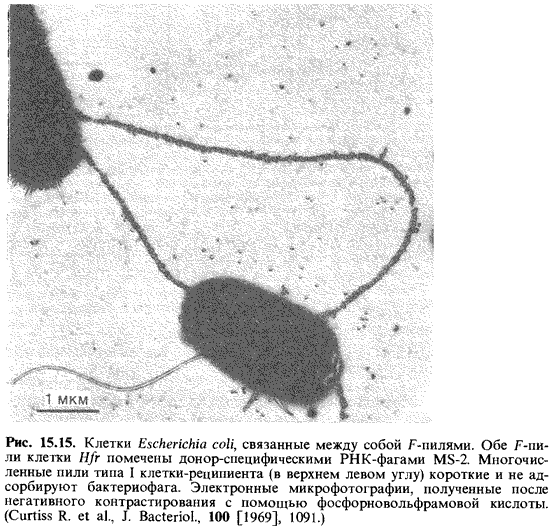
Фактор F представляет собой кольцевую двухцепочечную молекулу ДНК с массой 45 106 Да. В качестве внехромосомного автономно ре-плицируемого элемента ДНК ее следует отнести к плазмидам. Эта молекула содержит гены, ответственные за процесс конъюгации, в том числе гены, детерминирующие особые структуры клеточной поверхно сти, например половые волоски, или F-пили (рис. 15.15), необходимые для конъюгации. По всей вероятности, они служат для взаимного узна вания при контакте между клеткой-донором и клеткой-реципиентом и делают возможным образование конъюгационного мостика, по кото рому ДНК переходит внутрь клетки-реципиента. Пока не ясно, происхо дит ли такая «инъекция» ДНК через сами F-пили.
В популяции F+ лишь немногие клетки способны быть донорами хромосомной ДНК. Оказалось, что это те клетки, в которых фактор F интегрировался в бактериальную хромосому (рис. 15.16). Если клоны таких клеток-доноров использовать в экспериментах со скрещиванием, то рекомбинанты образуются примерно в тысячу раз чаще, чем при ис-

пользовании обычных клеток F +. Клетки-доноры, обеспечивающие вы сокую частоту рекомбинаций, получили название клеток Hfr (от англ. high frequency of recombinants). Фактор F включается в бактериальную хромосому лишь в определенных участках, число которых ограниченно; этот процесс сравним с интеграцией фага X (лямбда) в хромосому клет ки-хозяина (см. рис. 4.14, 15.12 и 15.16).
Процесс переноса. Если смешать популяцию клеток Hfr с избытком клеток F ~, то почти каждая клеткаHfr найдет себе партнера F ~ и бу дет с ним конъюгировать. Из такой смеси через определенные проме жутки времени брали пробы и, сильно встряхивая их в смесителе, на сильственно разъединяли партнеров. Затем пробы переносили на чашки с агаром для выделения рекомбинантов. И наконец, исследовали ре-комбинантные штаммы, чтобы выяснить, какие гены были переданы до норами клеткам-реципиентам. Исследования показали, что каждый ген передается в совершенно определенный момент времени после начала конъюгации (рис. 15.16). Временная последовательность переноса генов соответствовала порядку их расположения в бактериальной хромосоме, установленному в результате генетического анализа. Это значит, что любой штамм Hfr представляет собой гомогенную популяцию, все клетки которой передают свою хромосому реципиенту одинаковым образом-начиная с определенного участка (начала) и в одном и том же направлении. Чем дальше располагается тот или иной ген от «начала» хромосомы, тем позже он передается и тем реже попадает внутрь клет ки-реципиента, даже если конъюгацию не прерывать искусственно, Перенос всей хромосомы Е. coli продолжается при 37°С около 100 мин. Эксперименты, осуществленные по принципу «прерванной конъюгации», сделали возможным составление генетических карт.
Разные штаммы Hfr, выделенные независимо друг от друга из одно го и того же штамма F + , различаются по двум главным признакам: роль «начала» играет у каждого штамма иная точка хромосомы и каждый штамм отличается своей специфической последовательностью переноса генов. Результаты экспериментов согласуются с представле нием о том, что фактор F при интеграции (т. е. при переходе в состоя ние Hfr)может включаться в бактериальную хромосому в одном из примерно 20 возможных генных локусов. При переносе бактериальная ДНК реплицируется, начиная от места включения фактора F, и вновь синтезированная цепь, двигаясь 5'-концом вперед, проталкивается внутрь клетки-реципиента. Вслед за этим процессом переноса в клетке-реципиенте происходит гомологичная рекомбинация между донорской ДНК и собственной ДНК реципиента. Взаимоотношения между клет кой F~, клеткой F+ и клетками Hfr представлены на рис. 15.16.
Генетическая карта. В результате применения описанного выше ме тода прерванной конъюгации, позволяющего выяснить временную по следовательность переноса генов из клетки-донора, можно составить карту расположения генов в бактериальной хромосоме (рис. 15.17). Ско рость их переноса в течение всего процесса остается постоянной. Мо менты перехода внутрь клетки-реципиента позволяют судить о расстояниях между ними в хромосоме. При использовании этого метода не удается учитывать различия менее одной минуты. Для более тонкого картирования может служить анализ сцепления при трансдукции (переносе генов фагом).
Для Escherichia coli K12 известно расположение более чем тысячи ге нов, главным образом структурных, кодирующих ферменты. Последо вательность расположения генов на бактериальной хромосоме была определена для Salmonella typhimurium, Streptomyces coelicolor, Bacillus subtilis и некоторых других бактерий.
Перенос генов при посредстве фактора F'. Интеграция (включение) фактора F в бактериальную хромосому обратима. F-фактор может быть высвобожден из хромосомы, и тогда клетка Hfr становится клет кой F+ (рис. 15.16). Этот процесс «вырезания» (эксцизии, выключения) происходит примерно с той же частотой, что и интеграция. При пра вильной эксцизии разрыв происходит в том же самом месте, что и при интеграции. В редких случаях он происходит где-то очень близко к это му месту, и в результате соседний участок ДНК остается присоеди ненным к фактору F. Этот фактор F, содержащий небольшой фрагмент

хромосомной ДНК, называют фактором F'. Возникновение фактора F' аналогично образованию фага, осуществляющего специфическую транс-дукцию (разд. 15.3.3).
Клетку, содержащую фактор F', называют первичной клеткой F'. Включившаяся в F-фактор ДНК теперь может передаваться клетками-донорами F' штаммам F~ с такой же высокой частотой (100%), что и при обычной передаче фактора F штаммами F+ реципиентам F ~. Тот же самый фрагмент ДНК мог бы передаваться штаммом Я/г штамму F ~ с максимальной частотой 1%. Если фактор F' будет перене сен из первичной родительской клетки F' (где он впервые возник) в нор мальную клетку F ~, то образуется вторичная клетка F', в которой не большой участок бактериальной хромосомы окажется удвоенным (будет в диплоидном состоянии).
Распространенность конъюгации среди других групп бактерий. Переда ча генов путем конъюгации, открытая у Escherichia coli, очень широко распространена у энтеробактерий.
Посредством переноса факторов F из Е. coli K12 в клетки Salmonella и Shigella удалось создать новые генетические системы. Сходные си стемы были найдены в группе псевдомонад. Конъюгация у энтеробактерий представляет собой высокоразвитый процесс; чтобы она осуществи лась, достаточно суспендировать смесь клеток-партнеров в жидкой среде и оставить на некоторое время в покое. У многих других бакте рий конъюгацию удается вызвать лишь в том случае, если колонии обо их партнеров будут хорошо перемешаны и размазаны на твердой среде, где они должны затем расти несколько дней. Если теперь распределить клетки по одной на селективной среде, то окажется, что у многих кле ток возникла новая комбинация признаков и, по всей вероятности, про изошел обмен крупными участками бактериальных хромосом. Про цессы конъюгации широко изучались на Streptomyces coelicolor, видах Nocardia, Rhizobium и других бактериях. Обмен генами путем конъюга ции и мобилизация генов с помощью плазмид, вероятно, очень распространены в мире прокариот.
Плазмиды
Многие (если не все) бактерии могут содержать внехромосомные эле менты ДНК. Эти малые по сравнению с бактериальной хромосомой, замкнутые в кольцо двухцепочечные ДНК называют плазмидами. При росте в обычных условиях бактерии могут без них обходиться: клетки, «излеченные» от плазмид с помощью УФ-облучения, митомицина С или акридинового красителя, хорошо растут на обычных пита тельных средах. Плазмиды распознаются по особым свойствам, ко торые приобретает содержащая их клетка. Некоторые плазмиды де лают клетку способной конъюгировать с другими клетками. Это обеспечивает дальнейшее распространение таких плазмид путем прямо го межклеточного контакта. С прототипом подобного рода плазмиды мы уже встречались (с. 457) при рассмотрении F-факторов Escherichia coli.
Факторы фертильности (F-факторы). Это, как уже говорилось, плаз миды, которые могут включаться в бактериальную хромосому подобно ДНК умеренного фага лямбда. Они «мобилизуют» генетическую информацию этой хромосомы и осуществляют перенос ее в другую клет ку. Такой перенос (конъюгация) был хорошо изучен на Е. coli.
Факторы резистентности (R-факторы). Бактерии, устойчивые (рези стентные) к некоторым антибиотикам, были впервые открыты в 50-е годы в Японии. Речь идет о штаммах возбудителя дизентерии Shigella,выделенных от больных, которых лечили антибиотиками. Характерно то, что бактерии обнаруживали множественную устойчивость и что эта устойчивость могла передаваться другим бактериям, таким какEscherichia coli. Как стало теперь известно, факторы резистентности (R) содержат гены, которые делают клетку устойчивой, например, к сульфонамидам, стрептомицину, хлорамфениколу и тетрациклину. Некоторые К-факторы обусловливают резистентность сразу к восьми ан тибиотикам, другие же придают устойчивость к ядовитым тяжелым ме таллам, например ртути, никелю, кадмию или кобальту. К-плазмида несет две группы генов: 1) гены, ответственные за передачу плазмиды путем конъюгации (гены tra),-они образуют так называемый «фактор переноса устойчивости» (RTF, resistence transfer factor); 2) гены, обусло вливающие собственно резистентность (они составляют лишь неболь шую часть плазмиды) (рис. 15.18).
Фактор переноса устойчивости (RTF) включает все гены, ответ ственные за перенос фактора R из клетки в клетку, который осущест вляется обычно путем конъюгации. Таким образом, фактор R, так же как и фактор F, в широком смысле инфекционен. Область RTF по своей молекулярной структуре гомологична соответствующей области F-фак-тора Е. coli. Для некоторых К-факторов характерен широкий круг хо зяев; возможен их перенос между несколькими разными родами бакте рий, что способствует их дальнейшему распространению. В некоторых случаях наблюдали, что вместе с фактором R передаются и хромо сомные гены, которые, по-видимому, были мобилизованы им.
Механизм устойчивости к антибиотикам, определяемой К-факторами, может быть не таким, как в случае ее хромосомного наследования. На глядным примером этого служит резистентность к стрептомицину. Если она зависит от хромосомного гена, то она связана с изменением субъ единицы 30S рибосомы, так что бактерия не имеет мишени для воздей ствия стрептомицина (разд. 2.2.2). В отличие от этого устойчивость, обусловленная К-фактором, основана на инактивации антибиотика в ре зультате его аденилирования под влиянием фермента. Ферментативная химическая модификация антибиотиков часто бывает причиной устой чивости к ним, обусловленной плазмидами; например, хлорамфеникол ацетилируется, канамицин и неомицин подвергаются фосфорилирова-
нию и ацетилированию, а пенициллин инактивируется пенициллиназой. Поскольку и при наличии К-факторов возможна генетическая рекомби нация, может возникнуть новое сочетание генов, придающее дополнительные свойства устойчивости. К-факторы имеют большое значение для химиотерапии; их существование-лишний довод против бескон трольного применения антибиотиков, так как они могут распростра няться в популяциях бактерий подобно инфекционным агентам.
Бактериоцины. Многие бактерии синтезируют белки, убивающие родственные виды или штаммы или тормозящие их рост. Эти белки с весьма специфическим действием, бактериоцины, кодируются особыми плазмидами, бактериоциногенными факторами. Бактериоцины были вы делены из Escherichia coli(колицины), Pseudomonas aeruginosa (пиоцины), Bacillus megaterium (мегацины) и других бактерий.
Другие признаки, определяемые плазмидами. Плазмиды могут содер жать также гены, обусловливающие ряд специфических биологических свойств, которые в определенных условиях создают селективное пре имущество. Гены ферментов, необходимых для расщепления камфоры, салициловой кислоты, нафталина, октана, 2,4-дихлорфеноксиуксусной кислоты и многих других необычных субстратов, могут находиться в плазмидах. Мы уже упоминали о плазмиде бактерии Agrobacterium tumefaciens,вызывающей опухоли у растений, и ее биохимической актив ности (разд. 4.3). Перечень свойств, наследуемых с плазмидами, стал сейчас очень длинным и включает, в частности, азотфиксацию, образование клубеньков, синтез индолилуксусной кислоты, диацетила, гидроге-назы, поглощение Сахаров. Некоторые из этих свойств могут опреде ляться генами бактериальной хромосомы; это свидетельствует о том, что более или менее часто происходит обмен генами или группами ге нов между хромосомой и плазмидой. Плазмиды, вероятно, играли очень важную роль в эволюции прокариот.
Несовместимость. Многие бактерии содержат плазмиды различной величины. Сосуществование разных плазмид в одной бактериальной клетке говорит о том, что такие плазмиды совместимы между собой. Однако две родственные плазмиды не могут сосуществовать в одной клетке-они несовместимы. Все плазмиды подразделяются на группы несовместимости: плазмиды, относящиеся к одной и той же группе, несовместимы друг с другом.




