В настоящее время известны по меньшей мере три разных механизма рекомбинации попавшей в бактериальную клетку чужеродной ДНК с бактериальной хромосомой (или с плазмидой) in vivo: 1) общая гомо логичная рекомбинация, 2) сайт-специфическая рекомбинация и 3) него мологичная рекомбинация.
Общая гомологичная рекомбинация. В этом случае поступившая из вне ДНК рекомбинируется с клеточной ДНК путем реципрокного обме на соответствующими участками. Если не считать различий, обусло вленных мутациями, партнеры по рекомбинации должны иметь одинаковую нуклеотидную последовательность, т. е. быть максимально гомологичными. Гомологичная рекомбинация находится под контро лем гена гее А; мутанты с дефектом этого гена (гее ') не способны к го мологичной рекомбинации.
Существует несколько моделей данного механизма. Предполагают, что спаривание оснований происходит между деспирализованными, одноцепочечными участками двух двойных цепей ДНК. Вторая цепь, возможно, образуется в результате репликации или репарации.
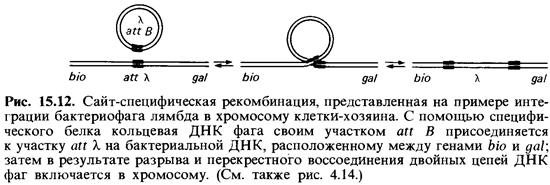
Сайт-специфическая рекомбинация. Этот процесс осуществляется не зависимо от гомологичной рекомбинации, т.е. возможен и у мутантов гее ~. Он состоит в том, что короткая двухцепочечная ДНК встраивает ся в определенном месте в длинную двойную спираль; при этом мень ший партнер теряет свою автономность. Типичным примером сайт-спе цифической рекомбинации может служить интеграция бактериофага лямбда (к) (рис. 4.14).
Генетические эксперименты свидетельствуют о том, что фаг при переходе в состояние профага включается в хромосому клетки-хозяина в определенном месте-между да/-опероном и биотиновой областью (рис. 15.12). Включению фага предшествует его присоединение к опреде ленному участку бактериальной ДНК. Ранее считали, что оно опреде ляется высокой степенью гомологии нуклеотидных последовательно стей, однако эта гомология оказалась незначительной; по-видимому, большую роль здесь играет кодируемый фагом белок-так называемая интеграза. В определенном участке фаговой ДНК (att В) и в соответствующем участке бактериальной ДНК (att X) этот белок катализирует разрыв и перекрестное воссоединение геномов фага и клетки-хозяина.
Негомологичная рекомбинация. Рекомбинационные процессы, в ко торых участвуют сегменты ДНК, не обнаруживающие заметной генети ческой гомологии, называют негомологичной рекомбинацией. Так же как и сайт-специфическая рекомбинация, она представляет собой инте грационную форму рекомбинации, т.е. не обмен, а соединение ДНК. Негомологичная рекомбинация независима от гена гее А. К такой ре комбинации способны: 1) вставочные последовательности (IS-эле-менты); 2) транспозоны (Тп); 3) бактериофаг (х (мю). Молекулярный ме ханизм негомологичной рекомбинации еще не вполне выяснен.
Как было установлено около 15 лет назад, некоторые мутации, спон танно возникающие у Escherichia coli,объясняются включением чуже родной ДНК. Такие мутации происходят в структурных и регуляторных генах по всей хромосоме. Чужеродная ДНК представляет собой так на зываемые инсерционные последовательности (IS-элементы); они встре чаются как в бактериальных хромосомах, так и в плазмидах. IS-эле менты состоят из 800-1400 пар нуклеотидов; распознаваемых феноти-пических признаков они не кодируют, и об их функциях мало что известно. Мутагенное действие их обусловлено просто включением по сторонней ДНК, нарушающим процесс транскрипции (с. 447). Можно предполагать, что IS-элементы играют важную роль в перестройках ге нетического материала.
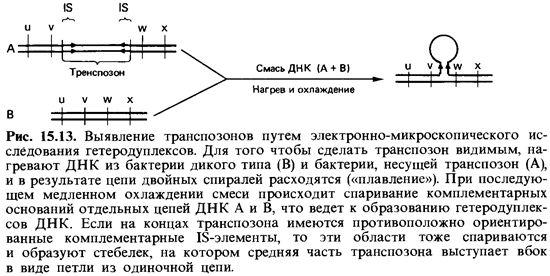
Транспозоны это последовательности ДНК, которые способны встраиваться во многие участки генома и могут «перепрыгивать» с плазмиды на бактериальную хромосому, на другую плазмиду или на умеренный фаг. Транспозоны содержат гены, определяющие внешне распознаваемые признаки, а именно устойчивость к таким антибиоти кам, как пенициллин, тетрациклин или канамицин. В связи с этим их легче обнаружить, чем IS-элементы. По обе стороны от генов устойчи вости, находящихся внутри транспозона, расположены две одинаковые последовательности, которые могут идти в одном и том же или в про тивоположных направлениях. Эти повторяющиеся последовательности оснований ДНК частью идентичны с IS-элементами. Расположение этих «фланкирующих» отрезков ДНК можно определить путем электронно-микроскопического исследования гетеродуплексов (рис. 15.13).
Бактериофаг мю сходен с IS-элементами и с транспозонами необычностью своего поведения при включении в бактериальную хромосому. Он обладает типичными свойствами фага, и в то же время его можно рассматривать как гигантский транспозон.




