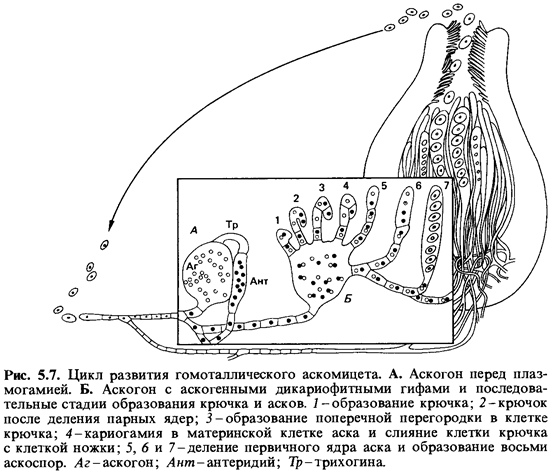
Цикл
развития (рис. 5.7). Выходящая
из аскоспоры ростовая трубка дает начало мицелию. У многих форм на мицелии
появляются конидиеносцы. Многочисленные конидии, прорастая, образуют мицелии,
сходные с теми, которые развиваются из аскоспор. На том же самом мицелии, где
формируются конидии, позднее появляются аски. Фаза полового размножения начинается
с образования аскогонов. В типичном случае аскогон несет особую трубочку - трихогину,
в которую попадают мужские ядра. Эти ядра через трихогину переходят из
антеридиев в аскогон (плазмогамия), где происходит спаривание ядер, но еще не
слияние. Затем из аскогона вырастают так называемые аскогенные гифы, клетки
которых содержат по одному мужскому и одному женскому ядру (дикариотические
гифы). Эти ядра одновременно делятся. Слиянию ядер дикариона предшествует
специальное деление клетки с образованием крючка: кончик гифы
крючкообразно искривляется, происходит сопряженное деление обоих ядер и верхняя
пара дочерних ядер отделяется поперечными перегородками как от клетки, лежащей
ниже изгиба, так и от крючка. Крючок сливается с клеткой, находящейся ниже
изгиба, и в результате снова образуется клетка с двумя ядрами. Верхняя клетка в
дальнейшем становится аском. В ней происходит слияние обоих ядер.
Образовавшееся первичное ядро аска (диплоидное) претерпевает три деления, одно
из которых - меиотическое; в результате получается восемь дочерних ядер и в
конечном счете - восемь аскоспор. Число делений может быть меньшим (в этом
случае образуются 4 споры) или большим (число спор иногда превышает 1000).
Таким образом, и аскоспоры, и мицелии гаплоидны.
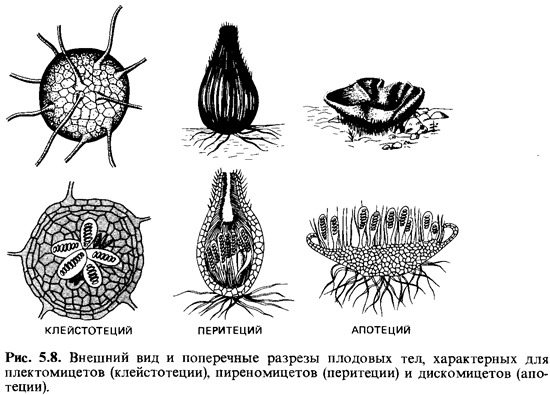
Плодовые
тела. Образование асков обычно
происходит в плодовых телах, или аскокарпах (впрочем, есть и исключения). Аскокарпы
- это вместилища, где созревают половые органы гриба. Сплетение гиф придает
плодовому телу характерный вид. Различают три формы плодовых тел (рис. 5.8): 1)
совершенно замкнутые плодовые тела, или клей стотеции, характерные для
плектомицетов; 2) бутылковидные плодовые тела - перитеции, типичные для
пиреномицетов; и наконец, 3) открытые чашеобразные плодовые тела - апотеции,
типичные для дискомицетов. Кроме того, существуют грибы с голыми асками
(протоаскомицеты). У трюфелей (Tuberales) аскокарпы остаются закрытыми.
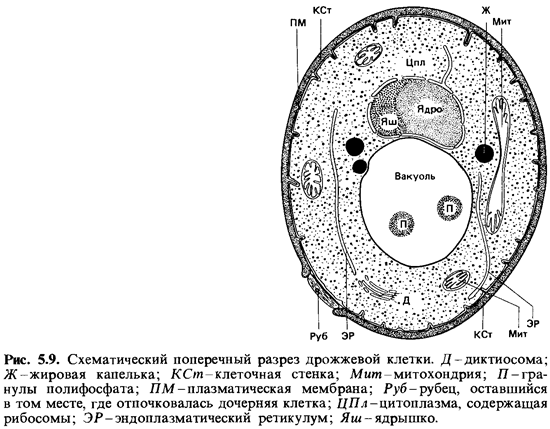
Дрожжи. К протоаскомицетам относятся дрожжи, или
почкующиеся грибы. Для дрожжей в широком смысле слова типичным способом бесполого
размножения является почкование (см. рис. 5.2); бинарное деление наблюдается
редко. Клетки, получающиеся в результате почкования, могут оставаться
соединенными, образуя псевдомицелий; в других случаях они расходятся. Аскоспоры
образуются в голом аске, развившемся из зиготы или из одной вегетативной
клетки.

Представители семейства Endomycetaceae размножаются
почкованием, но образуют также и мицелий. У Endomycopsis наряду с
гифами встречаются почкующиеся клетки и аски с аскоспорами. У Endomyces lactis (Geotrichum candidum,
Oospora lactis) гифы распадаются на артроспоры, т.е. на отдельные
самостоятельные клетки.
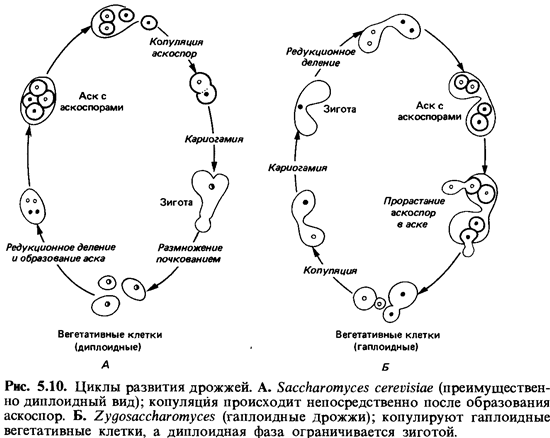
У Saccharomycetaceae, или
собственно дрожжей (рис. 5.9 и 5.10), мицелий отсутствует. Пекарские и пивные
дрожжи представляют собой физиологические расы Saccharomyces cerevisiae. Гаплоидные
почкующиеся клетки дрожжей могут сливаться (копулировать). За кариогамией может
сразу же следовать редукционное деление (мейоз) и образование четырех аскоспор.
Однако диплоидные клетки тоже способны размножаться почкованием; они крупнее и
физиологически активнее гаплоидных. В промышленности используют преимущественно
диплоидные и полиплоидные расы. Лишь в неблагоприятных условиях (например, на
среде с ацетатом) диплоидная клетка превращается в аск.
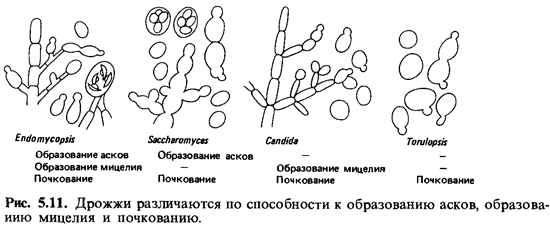
Еще более упрощенными формами следует считать
аспорогенные дрожжи. Только некоторые из них образуют мицелий; большинство
размножается исключительно почкованием. К аспорогенным дрожжам относятся роды Candida, Torulopsis, Cryptococcus, Rhodotorula,Pullularia и
ряд других (рис. 5.11).
Дрожжи встречаются в природе всюду, где есть
поддающиеся сбраживанию сахаристые соки: в нектаре цветков, на фруктах, на
листьях.
Pullularia pullulans принадлежит к группе дрожжей, вызывающих
образование черного налета на листьях, покрытых медвяной росой.
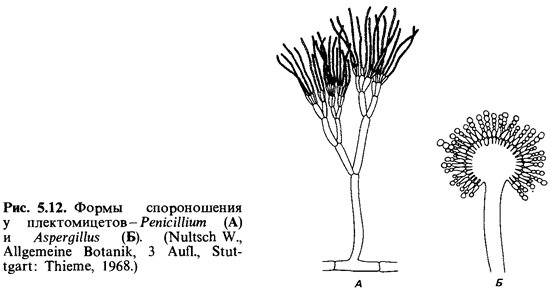
Плектомицеты. Плектомицеты - грибы, образующие
клейстотеции. К ним относятся такие важные роды, как Aspergillus (леечная
плесень) и Penicillium (кистевидная плесень), отличающиеся
друг от друга конидиальной стадией (рис. 5.12). О них мы еще не раз будем говорить
в связи с различными физиологическими вопросами. Сильно разветвленный
многоядерный мицелий плектомицетов усеян многочисленными конидиеносцами. При
развитии конидиеносца одна из клеток мицелия образует вырост, превращающийся в
вертикальную гифу. У Aspergillus такая гифа заканчивается
пузырьком, на поверхности которого вырастают стеригмы. На стеригмах образуются
конидии, располагающиеся как бусы на нитке. Конидии окрашены (они могут быть
черными, коричневыми, желтыми, зелеными и т. д.) и придают характерную окраску
всей колонии гриба.
Многие представители родов Aspergillus и Penicillium вызывают
порчу фруктов (при хранении), брезента, кожи и тому подобных материалов. Penicillium roqueforti и P. camemberti (теперь
их заменяют быстрее растущим P. caseicolum) придают сырам
«рокфор» и «камамбер» их специфический вкус. P. notatum и P.chrysogenum широко
известны как продуценты пенициллина.
Пиреномицеты. Плодовые тела у этих грибов - перитеции. В
типичных случаях перитеций имеет собственную (истинную) стенку. Аски, сидящие
на дне этого бутылковидного образования или на стенках его нижней части,
чередуются с особыми нитевидными выростами - парафизами. Канал шейки перитеция
выстлан перифизами (см. рис. 5.7 и 5.8).
К пиреномицетам принадлежит много вредоносных
грибов, в том числе облигатные паразиты - возбудители настоящей мучнистой росы (Erysiphe, Uncinula necator, Sphaerotheca mors-uvae и
др.), а также сапрофитные виды Chaetomium и виды Neurospora, известные
как объекты генетических исследований. Sordariafimicola и
некоторые другие виды - копрофилы. На пнях растут виды Xylaria и Hypoxylon с
черными конидиями, Nectaria galligena и N. cinnabarina вызывают
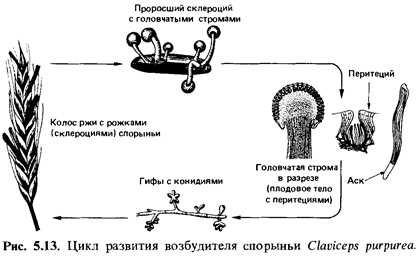
рак древесных пород. Claviceps purpurea - возбудитель
спорыньи (secale cornutum) ржи; другие виды Claviceps тоже
поражают различные злаки.
Цикл
развития пиреномицетов. Этот
цикл мы рассмотрим на примере Claviceps purpurea - спорыньи
ржи (рис. 5.13). Аскоспоры заражают во время цветения завязь ржи. Мицелий
разрастается в завязи и превращает ее в мягкую белую массу с желобками на
поверхности. В этих же лобках на тесно прижатых друг к другу кончиках гиф
образуется много мелких гиалиновых (бесцветных) конидий. Конидии, взвешенные в
сладком соке (медвяной росе), разносятся насекомыми. Вскоре все это образование
высыхает и пронизанная гифами завязь превращается в твердый, как рог,
склероций. Ко времени созревания злака склероций («маточные рожки») отпадают и
перезимовывают в почве. После периода покоя в условиях холода склероций весной,
при достаточной влажности и благоприятной температуре, прорастают, образуя
сидящие на ножках головки (головчатые стромы), в периферических слоях которых
находятся перитеции. В каждом аске Claviceps purpurea содержится
восемь нитевидных аскоспор.
Склероций содержат сильнодействующие
алкалоиды (производные лизергиновой кислоты - эргобазины, эрготоксины,
эрготамины), применяемые в качестве лечебных средств. Для получения этих
веществ рожь искусственно заражают спорыньей. Можно также получать глубинную
культуру одного из видов спорыньи (Claviceps paspali).
Дискомицеты. Дискомицеты образуют апотеции. К этой группе
принадлежат некоторые из более или менее известных лесных грибов,
например пецица (Peziza), сморчки (Morchella, Helvella) и
трюфели (Tuber), а также и вредные грибы, такие как Monilinia fructicola, Sclerotinia sclerotium, Rhytisma acerinumи Lophoderma (мы
назвали лишь некоторые). Апотеции - открытые плодовые тела, по форме напоминающие
блюдце. У некоторых лесных грибов они ярко-красные, желтые или оранжевые, у
других - черные или коричневые. На выжженной почве растет Pyronema omphalodes (P. confluens)
-излюбленный объект, на котором обычно демонстрируют образование асков.