Связывание М2-это
восстановительный процесс, и первым его продук том, который можно обнаружить,
является аммиак. Процесс восстано вления происходит на ферментном комплексе -
нитрогеназе. Нитрогена-за состоит из двух компонентов: белка, в состав которого
входят молибден, железо и сера, и белка, содержащего железо и серу.
Как сам фермент, так и процесс фиксации N2 отличаются крайней
чувствитель ностью к молекулярному кислороду. Это позволяет понять, почему как
у свободноживущих азотфиксирующих бактерий, так и в ткани клубень ков есть
особые механизмы, защищающие нитрогеназу от высокого парциального давления
кислорода.
Для
связывания молекулярного азота необходимы восстановитель ная сила и энергия
(рис. 13.2), которые могут быть получены в процессе фотосинтеза, брожения или
дыхания. В модельных экспериментах с очищенными компонентами нитрогеназной
системы (in vitro) можно доста влять энергию в виде АТР и восстановительную
силу в форме восстано вленных пиридиннуклеотидов и ферредоксинов, используя
переносчики, содержащие флаводоксин. Затраты АТР при этом очень высоки. 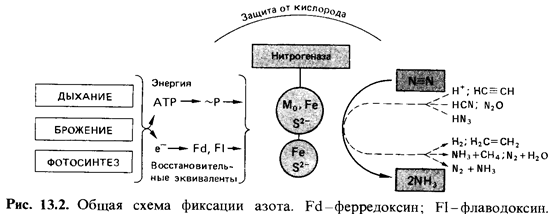
Нитрогеназная
система восстанавливает не только молекулярный азот (N=N), но и ацетилен
(НС=СН), азид, закись азота, цианид, ни триты, изонитрилы и протоны. На
восстановлении ацетилена основан наиболее простой метод, позволяющий выявить
нитрогеназу. Ацетилен восстанавливается только до этилена, который легко
поддается количе ственному определению с помощью газовой хроматографии. Все до
сих пор исследованные азотфиксирующие микроорганизмы и симбиотиче-ские системы
способны восстанавливать ацетилен.
Если не г
молекулярного азота, нитрогеназная система восстанавливает протоны до
молекулярного водорода. Таким образом, нитрогеназная система обладает также
свойствами АТР-зависимой Н2-образующей гидрогеназы. По скольку
молекулярный водород образуется и в присутствии N2, можно вклю чить
соответствующую реакцию в уравнение, описывающее фиксацию азота:
8[Н]
+ N2 + 2Н+
- 2NH4+ + Н2
Большинство
азотфиксирующих бактерий содержит наряду с нитрогеназой и (классическую)
гидрогеназу. активирующую Н2. Функция этой гидрогеназы за ключается,
по-видимому, в использовании водорода, образующегося при фикса ции
молекулярного азота.
Регуляция
связывания азота. У многих бактерий нитрогеназа обра зуется только
тогда, когда она необходима, т. е. в отсутствие подходя щего источника
связанного азота. Ионы аммония подавляют синтез ни-трогеназы. У пурпурных и
зеленых бактерий под влиянием этих ионов уменьшается также активность уже
синтезированного фермента. В регу ляции образования нитрогеназы большую роль,
очевидно, играет глу-таминсинтетаза. Глутаминсинтетаза и
глутаматсинтаза нужны бакте риям для включения ионов аммония в органические
соединения в том случае, если эти ионы присутствуют лишь в низкой концентрации.
Эта система обладает высоким сродством к ионам аммония и поддерживает их
концентрацию в клетке на низком уровне. Повышение концентрации ионов аммония в
окружении клетки (а тем самым и внутри клетки) по давляет образование
глутаминсинтетазы, а в результате-и нитроге назы.
Перенос
генов азотфиксацни («(/-генов). Способность к азотфиксации удается
передавать путем прямого межклеточного контакта от одной бактерии к другой.
Возможность передачи «(/-генов от Klebsiellapneumoniae к Escherichia coli путем
конъюгации, а также факт локализа ции этих генов в плазмиде позволяют
надеяться, что в близком буду щем удастся осуществить передачу их другим видам
бактерий1, а может быть, даже эукариотическим организмам. Но
поскольку для фиксации азота кроме нитрогеназы нужен еще специфический белок,
содержащий железо и серу, а также требуется защита этого фермента от 02,
по добные эксперименты сопряжены с большими трудностями. | 



