| Пятница, 03.05.2024, 00:12 | Главная | Регистрация | Вход |
|
Статистика

Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
Всего материалов в каталоге: 288
Показано материалов: 31-40 |
Страницы: « 1 2 3 4 5 6 ... 28 29 » |
Для понимания
структуры клеточной стенки бактерий важно прежде всего указать на сходство ее
опорного «скелета» с полимерами L-D-глюкозы
- целлюлозой и хитином.
Целлюлоза - главный
компонент клеточных стенок у высших растений, водорослей и оомицетов. У
бактерий целлюлоза в качестве мате риала клеточной стенки не встречается,
однако она соединяет клетки Sarcinaventriculi в большие
пакеты. Кроме того, целлюлозу выделяют Acetobacter aceti subsp. xylinum в
форме тонких фибрилл в среду, и она придает пленке «уксусного гриба»
("Mycoderma aceti") прочную кожистую консистенцию. |
У грам-положительных
бактерий доля муреиновой сетки составляет 30-70% сухой массы клеточной стенки
(толщиной в 40 слоев). Вместо м-диаминопимелиновой кислоты часто содержатся
LL-диаминопимелиновая кислота или лизин. У Staphylococcus aureus тетрапептидные
боковые цепи мурамовой кислоты связаны между собой межпептидными (например,
пентаглициновыми) цепочками. Участвующие в образовании таких структур
аминокислоты варьируют от вида к виду. Видоспецифическое строение опорного
каркаса представляет собой хороший таксономический признак. В клеточной стенке
грам-положительных бактерий полисахариды, если они вообще имеются, связаны
между собой ковалентно. Содержание белков невелико. Характерная особенность - наличие
тейхоевых кислот; это цепи, состоящие из 8-50 остатков глицерола или рибитола,
связанных между собой фосфатными мостиками. Некоторые из тейхоевых кислот
содержат эритритол или маннитол. Тейхоевые кислоты, вероятно, через фосфат
связаны с муреином по типу амида. |
У грам-отрицательных
бактерий муреиновая сеть однослойная и составляет менее 10% сухой
массы клеточной стенки (у Escherichia coli). Муреин содержит
только мезодиаминопимелиновую кислоту и не содержит лизина; межпептидные мостики
отсутствуют. Строение муреинового мешка у всех грам-отрицательных бактерий
одинаково. Наряду с этим опорным каркасом имеются большие количества
липопротеинов, липополисахаридов и других липидов, которые как бы наклеены
снаружи на муреиновый каркас. Они связаны ковалентно и составляют до 80% сухой
массы клеточной стенки. Для сохранения стабильности липополисахаридного слоя,
по-видимому, необходимы ионы кальция. У многих грам-отрицательных бактерий
муреиновый слой становится доступным для воздействия лизирующего муреин
фермента лизоцима только после обработки ЭДТА (этилендиаминтетраацетатом) для
удаления Са2 +. Этот комплексообразующий агент
вызывает освобождение части липополисахаридов. Тейхоевые кислоты у
грам-отрицательных бактерий до сих пор не были обнаружены. |
Структура
клеточной стенки и муреина была выяснена в основном в связи с изучением
действия лизоцима и пенициллина на бактерии. Открытый А. Флемингом (1922) лизоцим
является бактерицидным ферментом, содержащимся в слезной жидкости, в носовой
слизи и в яичном белке; лизоцим выделили также из бактерий (Escherichia coli, Streptomyces) и
бактериофагов. При воздействии лизоцима на суспензию грам-положительных
бактерий наблюдают быстрое ее просветление. Micrococcus luteus (lysodeikticus) лизируется
уже при концентрации 1 мкг лизоцима на 1 мл. Для лизиса клеток Bacillusmegaterium необходима
концентрация лизоцима 50 мкг/мл, а многие грам-отрицательные бактерии
растворяются толь ко тогда, когда к суспензии добавлен также
комплексообразующий агент (ЭДТА). |
Биосинтез и
встраивание структурных компонентов муреина в пептидогликановый скелет можно
разделить на три этапа (рис. 2.27). Первые стадии биосинтеза осуществляются в
цитоплазме. Здесь образуется пенталептид мурамовой кислоты. Синтез начинается с
образования N-ацетилглюкозамин-1-фосфата; в ряде последовательных
ферментативных реакций образуется лактиловый эфир, к которому присоединяются
пять аминокислот. Во время этого процесса растущая молекула остается связанной
с UDP (уридиндифосфатом), который служит носителем. 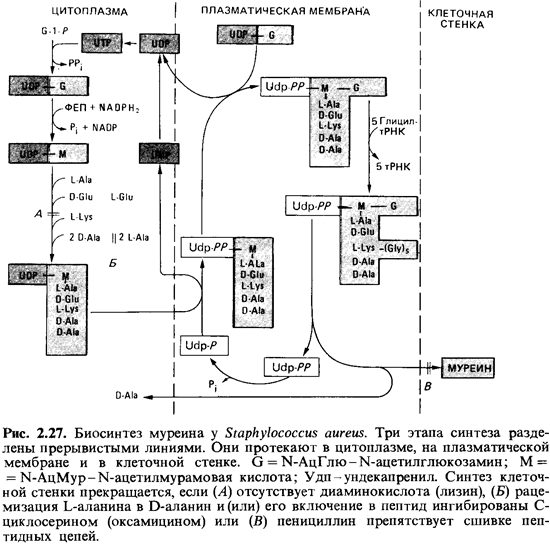
|
У этих
бактерий поверх однослойного или самое большее двуслойного муреинового мешка
располагается наружный слой клеточной стенки. На ультратонких срезах бактерий
он сходен по виду с плазматической мембраной - это так называемая наружная
мембрана. Этот слой стенки состоит из
белков, фосфолипидов и
липополисахаридов (рис. 2.28).
Со слоями
муреина - по-видимому, через диаминопимелиновую кислоту -ковалентно связаны
липопротеины; они ориентированы своими липофильными концами наружу и таким
образом закреплены в липофильном двойном слое (благодаря гидрофобному
взаимодействию). В этом слое находятся фосфолипиды и гидрофобные концы липополи
сахаридов; гидрофильные же
концы последних обращены наружу. |
Наружная
мембрана грам-отрицательных бактерий выполняет не только механические, но и
важные физиологические функции. В ее двойной липидный слой, состоящий из липида
А, полисахаридов и фосфолипидов, встроены белки, пронизывающие этот слой
насквозь. Вероятно, эти трансмембранные белки представляют собой заполненные
водой каналы - гидрофильные поры в липофильной мембране; поэтому их называют поринами. Существует
несколько различных поринов. Они пропускают через мембрану гидрофильные
низкомолекулярные вещества (вплоть до мол. массы порядка 6000). |
На клеточные
стенки многих бактерий наслаиваются снаружи более или менее толстые слои сильно
оводненного материала с высоким содержанием воды - капсулы и слизь. Такие
покровы не имеют для бактерий жизненно важного значения, но обладание капсулой
придает некоторым патогенным бактериям резистентность к фагоцитозу и тем самым
повышает их вирулентность для подопытных животных. |
Подвижность
бактерий может обеспечиваться различным образом. У большинства активно
передвигающихся, плавающих бактерий движение обусловлено вращением
жгутиков. Двигаться без жгутиков способны скользящие бактерии
(к которым относятся миксобактерии, цианобактерии и некоторые другие группы) и
спирохеты. О механизмах их движения будет сказано при рассмотрении
соответствующих групп бактерий. |
У многих
микроорганизмов в определенных условиях среды внутри клеток откладываются вещества,
которые можно рассматривать как запасные, - полисахариды, жиры, полифосфаты и
сера. Эти вещества накапливаются, если в питательной среде содержатся
соответствующие исходные соединения, но вместе с тем рост бактерий ограничен
или вообще невозможен из-за недостатка каких-то отдельных компонентов питания
или присутствия ингибиторов. Запасные вещества содержатся в клетках в
осмотически инертной форме - они нерастворимы в воде. При условиях,
благоприятных для роста, когда в этих веществах возникает потребность, они
снова включаются в метаболизм. Запасные полисахариды, нейтральные жиры и
поли-В-гидроксимасляная кислота могут служить источниками как энергии, так и
углерода. Поэтому при отсутствии внешних источников энергии они могут продлить
время существования клетки, а у спорообразующих видов - создать условия для
образования спор даже в отсутствие экзогенных субстратов. Полифосфаты могут
рассматриваться как резервный источник фосфата, а запасная сера - как
потенциальный донор электронов. |
|
 |  |


