Подвижность
бактерий может обеспечиваться различным образом. У большинства активно
передвигающихся, плавающих бактерий движение обусловлено вращением
жгутиков. Двигаться без жгутиков способны скользящие бактерии
(к которым относятся миксобактерии, цианобактерии и некоторые другие группы) и
спирохеты. О механизмах их движения будет сказано при рассмотрении
соответствующих групп бактерий. 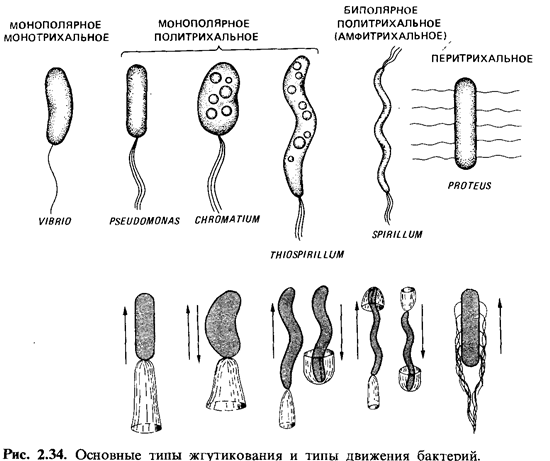
Расположение
жгутиков. Расположение жгутиков у подвижных эубактерий - это признак,
характерный для определенных групп, поэтому оно имеет таксономическое значение.
У палочковидных бактерий жгутики могут прикрепляться полярно или латерально (рис.
2.34). Среди бактерий с монополярным жгутикованием лишь немногие снабжены
только одним, но зато особенно толстым жгутиком - это монотрихи (Vibrio metschnikovii, рис.
2.35; Caulobacter sp.). У многих бактерий с монополярным и
биполярным жгутикованием одиночный по виду жгутик в действительности
представляет собой пучок из 2-50 жгутиков (политрихи).
Монополярно-политрихальное расположение жгутиков называют также лофотрихальным
(как у Pseudomonas, Chromatium), а биполярно-политрихальное
- амфитрихальным (у Spirillum). У Selenomonas имеется
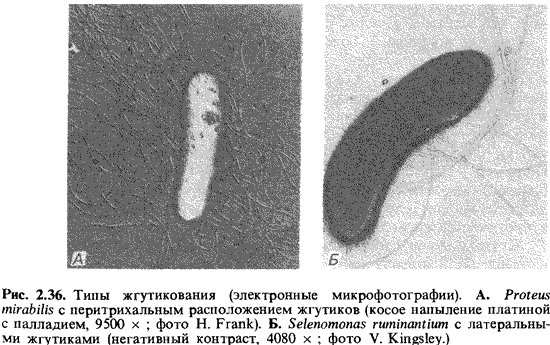
один пучок жгутиков, прикрепленный сбоку (рис. 2.36,2>). При перитрихальном
расположении (как у Enterobacteriaceae, Bacillaceae и не которых других
бактерий) жгутики располагаются по бокам клетки или на всей поверхности (рис.
2.36,4). 

Выявление
жгутиков. Рассмотреть жгутик (или пучок жгутиков) в проходящем свете
или в условиях фазового контраста удается только у немногих бактерий, например
у Chromatium okenii, Bdellovibrio,Thiospirillum (рис.
2.37). У многих других бактерий (Pseudomonas, Spirillum и
др.) жгутик и зону его биения можно увидеть только в темном поле. Легче всего
выявлять жгутики путем нанесения на них красителя или металла, а также с
помощью электронного микроскопа.
Функции
жгутиков. У большинства бактерий с полярным расположением жгутиков
последние действуют подобно корабельному винту и проталкивают клетку в
окружающей жидкой среде. Жгутик представляет собой спирально извитую нить,
приводимую во вращательное движение «мотором», находящимся в месте ее
прикрепления в плазматической мембране. Для перемещения клетки может служить
одиночный жгутик или пучок жгутиков. Жгутики вращаются сравнительно быстро;
например, у спирилл они совершают около 3000 оборотов в минуту, что близко к
скорости среднего электромотора. Вращение жгутиков приводит к тому, что тело
клетки вращается примерно с 1/3 этой скорости в противоположном направлении.
Жгутики могут
спонтанно или в ответ на внешний стимул изменять направление вращения (рис.
2.34). У некоторых бактерий с полярным расположением жгутиков это приводит к
тому, что клетка начинает двигаться вспять. Когда у Chromatium okenii в
ответ на вспышку света направление вращения жгутиков меняется, пучок жгутиков
превращается в тянущее приспособление; при этом назад клетка перемещается в
четыре раза медленнее, чем вперед, и ее движение становится «кувыркающимся». У Thiospirillum jenense
- гигантской фототрофной спириллы - единственный полярный пучок жгутиков
при обратном движении бьется уже не впереди клетки: пространство биения
жгутиков теперь охватывает клетку с боков: оно как бы вывернуто наизнанку
(подобно вывернутому ветром зонту). У спирилл с амфитрихальным расположением
жгутиков в таком положении находится, смотря по обстоятельствам, то один, то
другой пучок.
Перитрихально
расположенные жгутики Escherichia coli работают как один
хорошо скоординированный спиральный пучок и проталкивают клетку через среду. В
тех случаях, когда направление вращения отдельных жгутиков меняется, клетка
начинает «кувыркаться». По-видимому, перитрихально расположенные жгутики не
могут служить тянущим приспособлением.
Бактерии,
снабженные жгутиками, могут передвигаться очень быстро: Bacillus megaterium со
скоростью 1,6 мм/мин, Vibrio cholerae - 12 мм/мин. Это соответствует примерно от 300 до 3000 длин
тела в минуту.
Тонкое строение
жгутиков. Жгутики представляют собой спирально закрученные нити. У
разных бактерий они различаются по своей толщине (12-18 нм), длине (до 20 мкм),
а также по длине и амплитуде витка. Эти параметры характерны для каждого вида.
У некоторых бактерий могут образовываться жгутики разных типов. Нити жгутиков
состоят из специфического белка флагеллина. Они построены из субъединиц с
относительно малой молекулярной массой. Субьединицы располагаются по спирали
вокруг внутреннего свободного пространства (подобно белковым молекулам в вирусе
табачной мозаики). Таким образом, структура жгутика определяется свойствами
белковых субъединиц.
Жгутик
состоит из трех частей - описанной выше спиральной нити, «крюка» вблизи
поверхности клетки и базального тельца. С помощью базального тельца жгутик
закреплен в плазматической мембране и в клеточной стенке (рис. 2.38). Оно
состоит из центрального стержня, на котором у грам-отрицательных бактерий
находятся две пары колец. Наружная пара (кольца L и Р) расположены на уровне
наружного и внутреннего слоев клеточной стенки, а внутренняя пара (кольца S и
М) - на уровне наружного слоя плазматической мембраны. Так как у грам-положительных
бактерий наружная пара колец отсутствует, полагают, что для вращения жгутиков
необходима только внутренняя пара. Можно представить себе, что кольцо М
действует как приводной диск, а кольцо S играет роль подшипника на внутренней
поверхности пептидогликанового слоя. Молекулярный механизм вращательного
«мотора» жгутика пока не выяснен. 
О- и
Н-аитигены. Proteus vulgaris часто распространяется
по всей поверхности агара в виде тонкого серого налета (Н-форма, от нем. Hauch -
налет). Такое «роение» объясняется большой подвижностью клеток.
Некоторые штаммы налета не образуют (О-форма, от нем. ohne Hauch - без налета).
Эти штаммы неподвижны, они лишены жгутиков. Отсюда ведет свое начало обычная
терминология, принятая в бактериальной серодиагностике; антигены поверхности
или вообще тела клетки (соматические) называют О-антигенами, а антигены жгутиков
- Н-антигенами.
Фимбрии и
пили. Поверхность некоторых бактерий покрыта большим числом (от 10 до
нескольких тысяч) длинных, тонких прямых нитей толщиной 3-25 нм и длиной до 12
мкм, называемых фимбриями или пилями. Они встречаются как у жгутиконосных
видов, так и у форм, лишенных жгутиков. От них следует отличать половые пили,
или пили типа F, которые были обнаружены у клеток - доноров Escherichia coli К
12, т.е. у штаммов, содержащих половой фактор F (F +, Hfr).
Пили F встречаются только по одной или по две на клетку, они имеют вид полых
белковых трубочек длиной от 0,5 до 10 мкм.
Хемотаксис. Свободно
передвигающиеся бактерии способны к таксисам - направленным движениям,
определяемым внешними стимулами. В зависимости от факторов среды, вызывающих
направленное движение, говорят о хемотаксисе, аэротаксисе, фототаксисе и
магнитотаксисе.
Подвижные
бактерии реагируют на химические раздражители - скапливаются в одних местах, а
других мест избегают. Такая реакция свободно передвигающихся организмов
называется хемотаксисом. Скопления бактерий образуются под действием химических
факторов следующим образом (рис. 2.39). У форм с перитрихальными жгутиками
возможны только два типа двигательного поведения: прямолинейное движение и
кувыркание. Последнее прерывает прямолинейную пробежку и изменяет направление
пути. Когда бактерия оказывается в среде с градиентом концентрации
«привлекающего» ее субстрата (аттрактанта), ее прямолинейное движение длится
многие секунды, если она плывет по направлению к оптимальной его концентрации;
однако такое движение через несколько секунд прекратится, если бактерия плывет
в противоположном направлении. Хотя направление прямолинейного движения после
кувыркания оказывается совершенно случайным, тем не менее зависимость
длительности такого движения от его направления приводит в конечном результате
к накоплению бактерий в области оптимальной концентрации субстрата. За
чувствительность к химическому стимулу и за реагирование на него ответственны
хеморецепторы. В ряде случаев эти хеморецепторы действуют независимо от
способности бактерий утилизировать данный субстрат. Например, некоторые мутанты
продолжают совершенно нормально реагировать на определенное питательное
вещество, хотя и потеряли способность его использовать. 
Аэротаксис. У
подвижных бактерий можно определить тип метаболизма (аэробный или анаэробный)
по их аэротаксическим движениям и скоплению на определенных расстояниях от края
покровного стекла. В слое бактерий, помещенных между предметным и покровным
стеклами, аэрофильные бактерии скапливаются у края покровного стекла или в
непосредственной близости от оказавшихся в препарате пузырьков воздуха; это
указывает на их потребность в аэробных условиях и на то, что необходимую
энергию они получают за счет дыхания (рис. 2.40). Строго анаэробные бактерии
будут скапливаться в центре. Микроаэрофильные бактерии, например некоторые
псевдомонады и спириллы, будут держаться на определенном расстоянии от края. С
помощью бактерий, проявляющих положительный аэротаксис, Энгельману удалось
продемонстрировать выделение кислорода локально освещаемыми хлоропластами
зеленой водоросли Spirogyra. 
Фототаксис. Фототрофным пурпурным
бактериям для получения энергии необходим свет. Не удивительно поэтому, что в
результате фототаксиса они скапливаются в освещенном месте. Если выдержать в
темноте препарат, в котором плотная суспензия клеток Chromatium будет
равномерно распределена под покровным стеклом, а затем направить на него
сфокусированный пучок света, то бактерии сосредоточатся в области светового
пятна. Клетки, попавшие в это пятно случайно в результате своего беспорядочного
движения, уже не могут его покинуть. Как только они попадут в темную зону,
направление движения жгутиков мгновенно меняется на обратное и клетки
возвращаются в освещенное место. Изменение работы жгутиков происходит так
быстро, что эта реакция получила название «реакция испуга» (фоботаксис).
Впрочем, для того чтобы вызвать такой ответ, достаточно даже небольшого
различия в освещенности двух участков. Мелкие клетки Chromatium скапливаются
уже в таком месте, где освещенность всего на 0,7% выше, чем в окружающей
области. Таким образом, по своей чувствительности к световому контрасту они
приближаются к сетчатке человеческого глаза (для которой соответствующий порог
равен 0,4%).
Магиитотаксис. Из поверхностных слоев
донного ила пресноводных водоемов, а также морей были выделены бактерии
(палочки, спириллы, кокки), способные ориентироваться в магнитном поле и
перемещаться в направлении линий магнитного поля. Они содержат много железа
(0,4% сухого вещества) в форме ферромагнитной окиси железа (магнетита), которая
находится в гранулах (магнитосомах), расположенных около мест прикрепления
жгутиков. Бактерии, выделенные в северном полушарии, «ищут» север; здесь линии
магнитного поля проходят под углом около 70° к горизонту вниз, вглубь водоема.
Магнитотаксическоё поведение направляет бактерии в глубину ила, где очень мало
или вовсе нет кислорода. Так как магнитотаксические бактерии - анаэробы или микроаэрофилы,
их реакция на магнитное поле понятна с точки зрения экологии. Такие клетки,
завезенные в южное полушарие, в массе своей, конечно, погибнут; выживут лишь
немногие «неправильно» поляризованные клетки, которые могут затем
размножиться. Полярность, очевидно,
генетически не зафиксирована.
| 



