| Воскресенье, 10.05.2026, 00:53 | Главная | Регистрация | Вход |
|
Статистика
Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В категории материалов: 29
Показано материалов: 21-29 |
Страницы: « 1 2 3 |
Сортировать по:
Дате ·
Названию ·
Комментариям ·
Просмотрам
Биосинтез и
встраивание структурных компонентов муреина в пептидогликановый скелет можно
разделить на три этапа (рис. 2.27). Первые стадии биосинтеза осуществляются в
цитоплазме. Здесь образуется пенталептид мурамовой кислоты. Синтез начинается с
образования N-ацетилглюкозамин-1-фосфата; в ряде последовательных
ферментативных реакций образуется лактиловый эфир, к которому присоединяются
пять аминокислот. Во время этого процесса растущая молекула остается связанной
с UDP (уридиндифосфатом), который служит носителем. 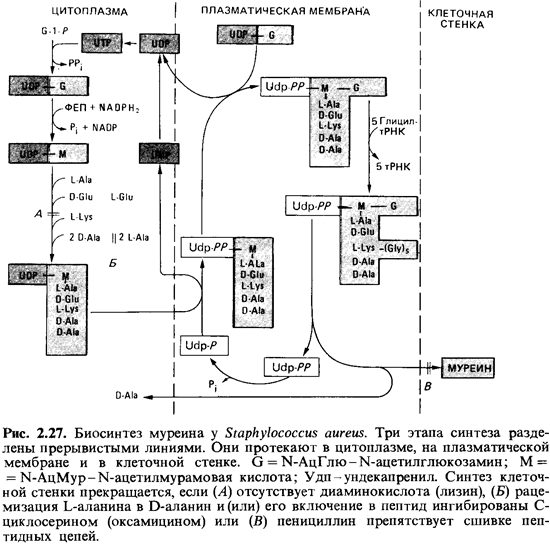
|
У этих
бактерий поверх однослойного или самое большее двуслойного муреинового мешка
располагается наружный слой клеточной стенки. На ультратонких срезах бактерий
он сходен по виду с плазматической мембраной - это так называемая наружная
мембрана. Этот слой стенки состоит из
белков, фосфолипидов и
липополисахаридов (рис. 2.28).
Со слоями
муреина - по-видимому, через диаминопимелиновую кислоту -ковалентно связаны
липопротеины; они ориентированы своими липофильными концами наружу и таким
образом закреплены в липофильном двойном слое (благодаря гидрофобному
взаимодействию). В этом слое находятся фосфолипиды и гидрофобные концы липополи
сахаридов; гидрофильные же
концы последних обращены наружу. |
Наружная
мембрана грам-отрицательных бактерий выполняет не только механические, но и
важные физиологические функции. В ее двойной липидный слой, состоящий из липида
А, полисахаридов и фосфолипидов, встроены белки, пронизывающие этот слой
насквозь. Вероятно, эти трансмембранные белки представляют собой заполненные
водой каналы - гидрофильные поры в липофильной мембране; поэтому их называют поринами. Существует
несколько различных поринов. Они пропускают через мембрану гидрофильные
низкомолекулярные вещества (вплоть до мол. массы порядка 6000). |
На клеточные
стенки многих бактерий наслаиваются снаружи более или менее толстые слои сильно
оводненного материала с высоким содержанием воды - капсулы и слизь. Такие
покровы не имеют для бактерий жизненно важного значения, но обладание капсулой
придает некоторым патогенным бактериям резистентность к фагоцитозу и тем самым
повышает их вирулентность для подопытных животных. |
Подвижность
бактерий может обеспечиваться различным образом. У большинства активно
передвигающихся, плавающих бактерий движение обусловлено вращением
жгутиков. Двигаться без жгутиков способны скользящие бактерии
(к которым относятся миксобактерии, цианобактерии и некоторые другие группы) и
спирохеты. О механизмах их движения будет сказано при рассмотрении
соответствующих групп бактерий. |
У многих
микроорганизмов в определенных условиях среды внутри клеток откладываются вещества,
которые можно рассматривать как запасные, - полисахариды, жиры, полифосфаты и
сера. Эти вещества накапливаются, если в питательной среде содержатся
соответствующие исходные соединения, но вместе с тем рост бактерий ограничен
или вообще невозможен из-за недостатка каких-то отдельных компонентов питания
или присутствия ингибиторов. Запасные вещества содержатся в клетках в
осмотически инертной форме - они нерастворимы в воде. При условиях,
благоприятных для роста, когда в этих веществах возникает потребность, они
снова включаются в метаболизм. Запасные полисахариды, нейтральные жиры и
поли-В-гидроксимасляная кислота могут служить источниками как энергии, так и
углерода. Поэтому при отсутствии внешних источников энергии они могут продлить
время существования клетки, а у спорообразующих видов - создать условия для
образования спор даже в отсутствие экзогенных субстратов. Полифосфаты могут
рассматриваться как резервный источник фосфата, а запасная сера - как
потенциальный донор электронов. |
К образованию
эндоспор способна только небольшая группа бактерий. Огромное значение эндоспор
связано с их термоустойчивостью. В то время как почти все остальные бактерии, а
также вегетативные клетки спорообразующих видов гибнут при 80°С (при температуре
пастеризации) через 10 мин, терморезистентные эндоспоры переносят
значительно более сильный нагрев; некоторые споры выдерживают даже кипячение на
протяжении многих часов. Трудоемкая и дорогостоящая техника стерилизации
(обеззараживания) рассчитана на уничтожение эндоспор. С другой стороны,
терморезистентность спор предоставляет своеобразную возможность избирательного
обогащения культур спорообразующих форм. Землю или иной материал, взятый в
определенном месте, на протяжении 10 мин прогревают при 80 или 100°С, что
приводит к гибели вегетативных клеток; только терморезистентные споры остаются
жизнеспособными и прорастают в подходящей питательной среде. |
Колониям
многих бактерий и грибов свойственна яркая окраска, обусловленная выделением
окрашенного продукта в окружающую среду или же пигментацией самой клетки.
Способность к образованию пигментов детерминирована генетически и поэтому может
использоваться в качестве характерного признака. Окрашенные формы легче
выявлять и идентифицировать. Среди пигментов могут встречаться представители
различных классов веществ: каротиноиды, феназиновые красители, пирролы,
азахиноны, антоцианы и др. |
Теория эндосимбиотического происхождения хлоропластов впервые была предложена в 1883 году Андреасом Шимпером, показавшим их саморепликацию внутри клетки. Её возникновению предшествовал вывод А.С. Фаминцина и О.В. Баранецкого о двойственой природе лишайников — симбиотического комплекса гриба и водоросли (1867 год). К.С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришел к выводу, что хлоропласты являются симбионтами, как и в случае с водорослями лишайника. В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов. |
|
 |  |


