В соответствии со своей ролью и функцией в балансе природы организмы разделяются на три группы. Зеленые растения синтезируют органические вещества, используя энергию солнца и углекислоту, поэтому их называют продуцентами. Животные являются потребителями (консументами); они расходуют значительную часть первичной биомассы для построения своего тела. Тела животных и растений в конце концов подвергаются разложению, при котором органические вещества превращаются в минеральные, неорганические соединения. Этот процесс, называемый минерализацией, осуществляют в первую очередь грибы и бактерии; в балансе природы они служат деструкторами. Таким образом, биоэлементы участвуют в циклических процессах. Здесь уместно коротко остановиться на биогеохимических круговоротах углерода, азота, фосфора и серы.
Круговорот
углерода. В круговороте
углерода микроорганизмы выполняют функцию, очень важную для поддержания жизни
на Земле. Они обеспечивают минерализацию углерода, переведенного зелеными
растениями в органические соединения, и тем самым поддерживают весьма
неустойчивое равновесие (рис. 1.1). Атмосферный воздух содержит чуть больше
0,03% двуокиси углерода (12 мкМ/л). 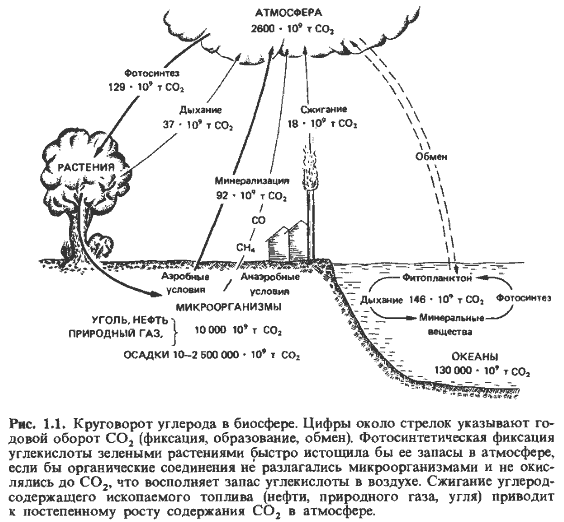
Фотосинтетическая же продуктивность зеленых
растений так велика, что запас СО2 в атмосфере был бы исчерпал
примерно за 20 лет. Это относительно короткий срок в человеческих масштабах
времени; ведь считается, что запасов энергии и угля на Земле хватит на срок от
1000 до 3000 лет. Да же если учесть запасы СО2 в океанах, то
этого газа хватило бы лишь примерно на 2000 лет.
Зеленым
растениям пришлось бы вскоре прекратить фиксацию СО2, если бы низшие
животные и микроорганизмы не обеспечивали возвращение этого газа в атмосферу в
результате непрерывной минерализации органического материала. В общем балансе веществ на
земном шаре почвенным бактериям и грибам принадлежит не меньшая роль, чем фотосинтезирующим
зеленым растениям. Взаимозависимость всех живых существ на Земле находит наиболее
яркое выражение в круговороте углерода.
Следует
отметить еще одну особенность процесса минерализации: небольшая часть
минерализованного углерода (1-1,5%) поступает в атмосферу не в виде СО2,
а в форме метана. Этот газ образуется из органических веществ в местах,
недоступных для кислорода воздуха (в почве тундр, на рисовых полях, в рубце
жвачных), попадает затем в атмосферу и окисляется там ОН-радикалами через окись
углерода (СО) до СО2. В процессе образования метана, так же как и
других газов, встречающихся в следовых концентрациях (Н2, СО, N2О,
NО2), участвуют главным образом бактерии.
Моря на
первый взгляд кажутся огромным резервом углекислоты. Однако следует учесть, что
скорость обмена СО2 атмосферы с С02 морей, где
более 90% этого вещества находится в форме HCOJ, очень мала; за один год таким
образом обменивается только десятая часть атмосферной двуокиси углерода. К тому
же в газообмене моря с атмосферой участвует лишь тонкий поверхностный слой
воды. Огромные количества С02, находящиеся в океанах ниже слоя
температурного скачка, выходят на поверхность лишь в немногих областях (Западная
Африка, Чили) и обогащают там атмосферу (до 0,05%). Уже на протяжении многих
лет содержание двуокиси углерода в воздухе неуклонно возрастает. С одной
стороны, это следует отнести за счет сжигания нефти и угля; в 1976 г. на Земле было
израсходовано (в основном сожжено) около 3,2*109 т нефти. С
другой стороны, повышение концентрации СО2 в атмосфере связано,
вероятно, с уменьшением фотосинтетической фиксации углерода в результате
сведения больших лесных массивов и деградации почвы. Следует подчеркнуть, что
Мировой океан представляет собой мощную буферную систему, которая стремится
поддерживать содержание СО2 в атмосфере на определенном уровне.
В результате
фотосинтетической фиксации СО2 зелеными растения ми образуются
в первую очередь сахара и другие родственные им со единения. Основная масса
фиксированного углерода как у древесных, так и у травянистых растений на время
откладывается в форме поли мерных углеводов. Примерно 60% двуокиси углерода,
фиксируемой на суше, идет на образование древесины. Древесина состоит на 75% из
полисахаридов (целлюлоза, гемицеллюлозы, крахмал, пектины и арабиногалактаны) и
содержит лишь немногим больше 20% лигнина и лигнанов; белка в ней очень мало
(1%). У злаков и других травянистых растений содержание полисахаридов еще выше.
Преобладание
полисахаридов среди продуктов ассимиляции зеленых растений обусловливает
большую роль Сахаров в питании всех живых организмов, нуждающихся в органической
пище. Глюкоза и другие сахара в форме полимеров - это количественно
преобладающие субстраты для процессов минерализации в природе; в виде мономеров
они служат предпочитаемыми питательными веществами для большинства гетеротрофных
микроорганизмов.
Круговорот
азота (рис. 1.2). Центральное место в круговороте азота занимает аммоний. Он
является продуктом разложения белков и аминокислот, попадающих вместе с
остатками животного и растительного происхождения в почву. В хорошо аэрируемых
почвах аммоний подвергается нитрификации; бактерии родовNitrosomonas и Nitrobacter окисляют
его до нитрита и нитрата. В качестве источника азота растения могут
использовать и ассимилировать как аммоний, так и нитрат. В отсутствие кислорода
из нитрата образуется молекулярный азот (денитрификация). Бактерии, участвующие
в этом процессе, используют при этом нитрат в качестве окислителя (акцептора
водорода), т.е. «дышат» с помощью NO2 вместо О2; в этом
случае говорят о «нитратном дыхании».
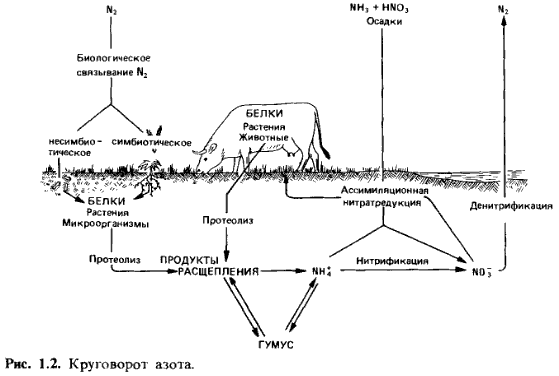
Денитрификация
ведет к потере азота почвой. Наряду с этим бактерии способны и к фиксации
молекулярного азота. Связывающие азот бактерии живут или свободно в почве (вне
симбиоза), или в симбиозе с высшими растениями (симбиотические азотфиксаторы).
Основную роль в круговороте азота наряду с животными и растениями играют
бактерии.
Круговорот фосфора. В биосфере фосфор
представлен почти исключительно в виде фосфатов. В живых организмах фосфорная
кислота существует в форме эфиров. После отмирания клеток эти эфиры быстро
разлагаются, что ведет к освобождению ионов фосфорной кислоты. Доступной для
растений формой фосфора в почве служат свободные ионы ортофосфорной кислоты (Н3Р04).
Их концентрация часто очень низка; рост растений, как правило, лимитируется не
общим недостатком фосфата, а образованием малорастворимых его соединений, таких
как апатит и комплексы с тяжелыми металлами. Запасы фосфатов в месторождениях,
пригодных для разработки, велики, и в обозримом будущем производство
сельскохозяйственной продукции не будет ограничиваться недостатком фосфора;
однако фосфат должен быть переведен в растворимую форму. Во многих местах
фосфат из удобрений попадает в проточные водоемы и озера. Так как концентрация
ионов железа, кальция и алюминия в водоемах невысока, фосфат остается в
растворенной форме, что приводит к эвтрофизации водоемов, особенно
благоприятной для развития азотфиксирующих цианобактерий. В почвах же из-за
образования нерастворимых солей фосфаты чаще всего быстро становятся
недоступными для усвоения.
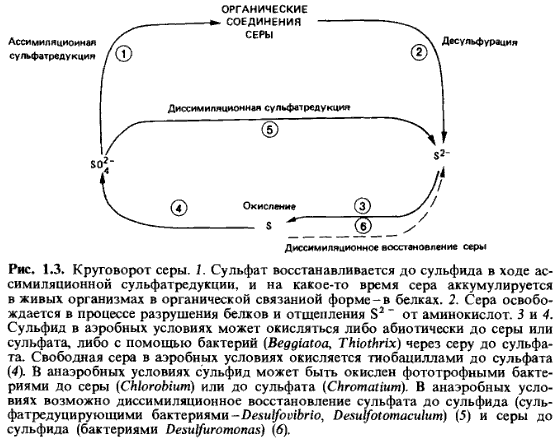
Круговорот
серы (рис. 1.3). В живых
клетках сера представлена главным образом сульфгидрильными группами в
серусодержащих аминокислотах (цистеин, метионин, гомоцистеин). В сухом веществе
организмов доля серы составляет 1%. При анаэробном разложении органических
веществ сульфгидрильные группы отщепляются десульфуразами; образование
сероводорода при минерализации в анаэробных условиях называют также
десульфурированием. Наибольшие количества встречающегося в природе сероводорода
образуются, однако, при диссимиляционном восстановлении сульфатов,
осуществляемом сульфатредуцирующими бактериями.
Этот
сероводород, образующийся в отсутствие молекулярного кислорода в осадках
водоемов, может быть окислен анаэробными фототрофными бактериями (Chromatiaceae)
до серы и сульфата. Когда сероводород
проникает в зоны, содержащие О2,
он окисляется либо абиотическим образом, либо аэробными
серобактерия ми до сульфата. Серу, необходимую для синтеза серусодержащих
аминокислот, растения и часть микроорганизмов получают путем ассимиляционной
сульфатредукции; животные же получают восстановленные соединения серы с пищей.
 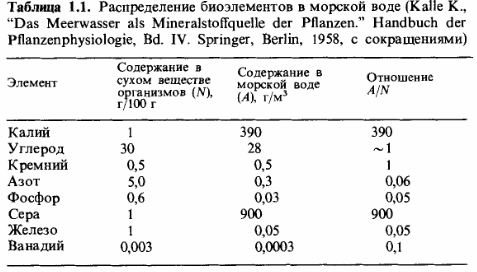
Фосфор и
азот как факторы, лимитирующие продукцию биомассы. Элементами, которые
ограничивают рост растений и тем самым продукцию биомассы, являются фосфор и
азот. Они играют такую роль и на суше, и в океанах. На этот счет имеются точные
данные для морской воды. Основываясь на данных табл. 1.1, можно рассчитать,
сколько биомассы может быть создано (в граммах сухого вещества) из элементов,
содержащихся в одном кубометре морской воды. Из 28 г углерода (С) может
образоваться 60-100 г
биомассы, из 0,3 г
азота (N) - 6 г,
а из 0,03 г
фосфора (Р) - только 5 г.
Отсюда следует, что продукцию биомассы в конечном счете лимитируют фосфаты.
Таким образом, в морской воде даже азотфиксирующие организмы – цианобактерии - не
имеют селективного преимущества перед другими. | 



