На электронных
микрофотографиях ультратонких срезов бактерий, фиксированных четырехокисью
осмия, плазматическая мембрана представляется многослойной. Она состоит из двух
осмиофильных и потому темных слоев толщиной 2-3 нм каждый и промежуточного
более светлого слоя толщиной 4-5 нм. По своему строению мембраны бактериальных,
животных и растительных клеток очень сходны. Это дает основание говорить об
универсальной «элементарной мембране» ("unit membrane").
Мембраны
можно выделить, подвергнув осмотическому шоку протопласты, полученные с помощью
лизоцима. Мембрана богата липидами, в особенности фосфолипидами
(табл. 2.1). Составляя всего 8-15% сухого вещества клетки, мембраны содержат
70-90% всех ее липидов. 
Плазматическая
мембрана состоит из двойного липидного слоя. Гидрофобные концы молекул
фосфолипидов и триглицеридов направлены внутрь, а гидрофильные «головки» -
наружу. Благодаря гидрофобным взаимодействиям между остатками жирных кислот,
входящих в состав липидов, и электростатическому взаимодействию между
гидрофильными «головками» мембрана стабилизируется. В двойной слой липидов
встроены белки - так называемые интегральные белки мембран. Они «плавают» в
этом слое, будучи погружены в него частично, или же пронизывают его насквозь.
Другие белки прикреплены к поверхности мембраны, и их называют периферийными
белками (рис. 2.20). Некоторые мембраны, по-видимому, с одной или с обеих
сторон покрыты сетью вытянутых белковых молекул. 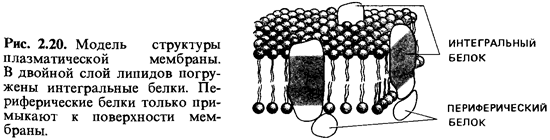
Мембрану
следует представлять себе как очень мягкое, пластичное, почти жидкое
образование; изолированные мембраны стремятся образовать замкнутые со всех
сторон пузырьки (везикулы); кусочки мембран сливаются краями друг с другом.
Плазматическая
мембрана играет важнейшую роль в обмене веществ. Она служит осмотическим
барьером клетки и контролирует как поступление веществ внутрь клетки, так и
выход их наружу. В мембране имеются механизмы активного транспорта и системы
субстрат - специфичных пермеаз. По-видимому, липидная пленка элементарной мембраны
пронизана «мостиками» (или каналами) из белков, и именно эти белки служат
порами, через которые осуществляется регулируемый транспорт веществ.
Ферменты
переноса электронов и окислительного фосфорилирования, находящиеся у эукариот в
митохондриях, у бактерий локализуются внутри или на поверхности плазматической
мембраны. Цитохромы, железо-серные белки и другие компоненты
электрон-транспортной цепи находятся исключительно в мембранах. Как показало
детальное изучение локализации отдельных компонентов, мембрана построена
асимметрично: например, цитохром с расположен в ее наружном слое, а
АТР-синтаза - на внутренней стороне мембраны. Мембрана характеризуется
направленным обменом веществ. По-видимому, мембраны ответственны и за другие
биосинтетические процессы, такие как синтез компонентов клеточной стенки и
капсулы, а также за выведение внеклеточных ферментов. Наконец, весьма
вероятно, что на мембране локализуется центр репликации РНК. Жгутики тоже
прикреплены к плазматической мембране.
Внутриклеточные
мембраны и ламеллы. У некоторых бактерий мембрана охватывает цитоплазму без
складок и впячиваний. У других она образует впячивания, пронизывает цитоплазму
или формирует мембранные тельца. У ряда бактерий описаны мезосомы (рис. 2.21).
Правда, в этом случае, вероятно, речь идет об артефактах, связанных с приготовлением
препаратов. 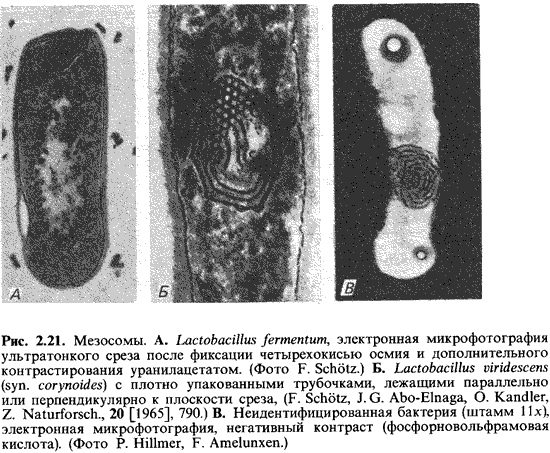
У Nitrobacter, Nitrosomonas и Nitrosococcus имеются
стопки ламелл, состоящие из параллельно расположенных плоских пузырьков, часть
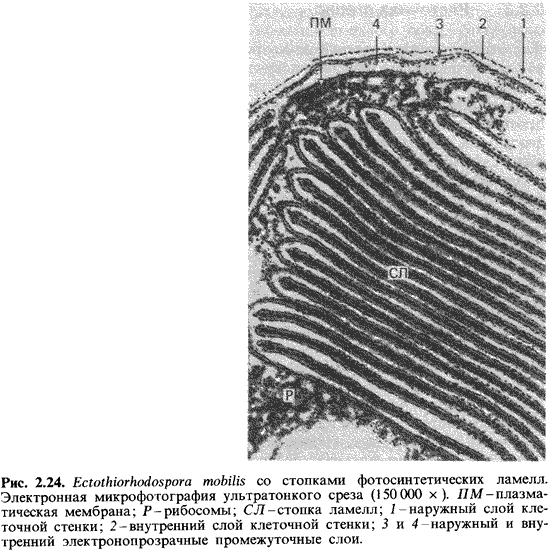
которых связана с плазматической мембраной (рис. 2.22 и 2.24).
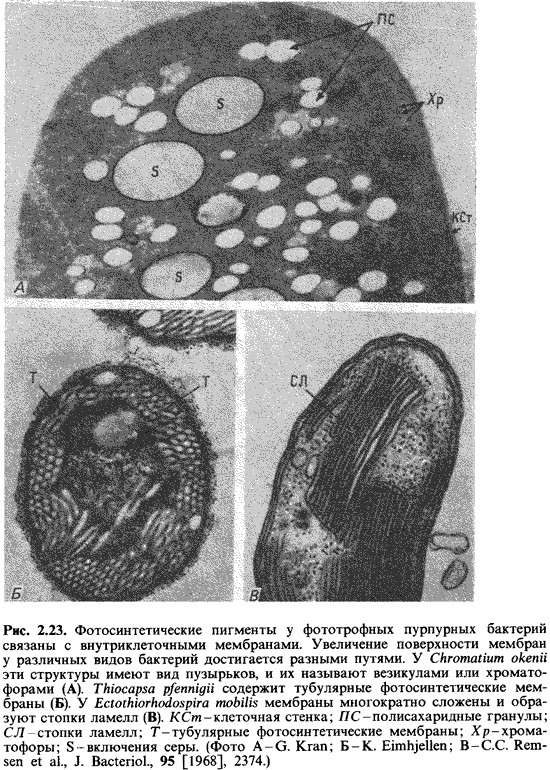
Особенно
богаты внутриклеточными мембранами фототрофные пурпурные бактерии. На
ультратонких срезах фотосинтетические внутренние мембранные системы имеют у них
вид трубок, пузырьков или стопок. У Rhodospirillum rubrum или
Chromatium клетка почти целиком заполнена пузырьками, которые
упакованы в округлые образования (рис. 2.23). Эти пузырьки, по-видимому,
возникают путем впячивания (инвагинации) плазматической мембраны и роста ее в
форме рукава. На этом рукаве на равных расстояниях образуются перетяжки, и в
резуль тате получаются пузырьки, не отделенные полностью друг от друга. Только
при разрушении и гомогенизации клеток эти пузырьки освобож даются, принимая вид
изолированных структур, которые раньше иногда называли «хроматофорами». У
других пурпурных бактерий пузырьки сильно уплощены и образуют упорядоченные
стопки (рис. 2.23, 2.24); последние по аналогии со структурой хлоропластов
зеленых растений называют стопками тилакоидов. 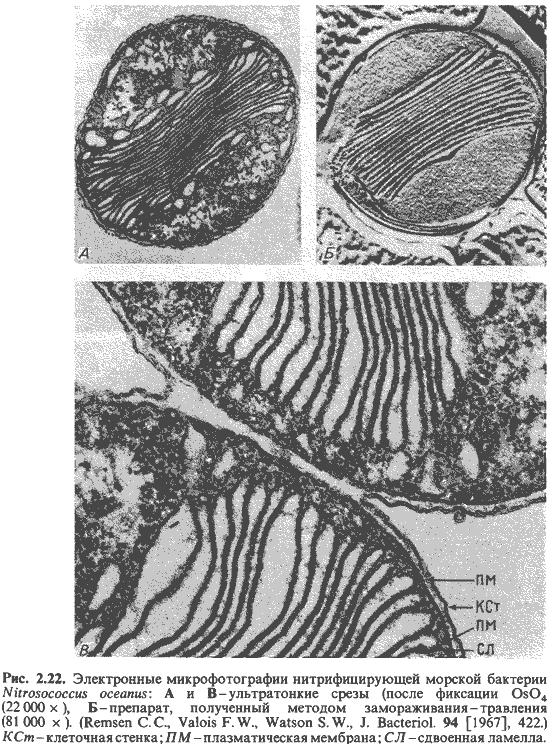
Фотосинтетические
мембраны сходны по строению и химическому составу с плазматической мембраной
(табл. 2.1), но, кроме того, содержат пигменты, поглощающие свет
(бактериохлорофиллы и каротиноиды), а также компоненты фотосинтетической
электрон-транспортной цепи (цитохромы, убихинон) и фосфорилирующей системы. | 



