Основываясь
на способе размножения - бинарном делении, цианобактерии уже давно объединили
вместе с бактериями под названием Schizophyta. Из-за общих физиологических
особенностей их, однако, относили к растениям и называли сине-зелеными
водорослями. Поэтому для них были действительны правила ботанической
номенклатуры. Только с тех пор, как научились ясно отличать прокариот от
эукариот, эту группу организмов причисляют к бактериям.
С водорослями
и высшими растениями цианобактерии имеют то общее, что все они осуществляют
фотосинтез с выделением кислорода и содержат хлорофилл а, а
также ряд других общих с растениями пигментов. Поэтому их и отнесли к
водорослям под названием сине-зеленых водорослей. Но уже Ф. Кон, учитывая
способ их деления, назвал их Schizophyceae и объединил со Schizomycetes
(бактериями), сведя в группу высшего порядка Schizophyta. Действительно, если
судить по строению клетки, наличию муреиновой клеточной стенки, 70 S-рибосомам
и другим определяющим признакам, приходится отнести их к грам-отрицательным
прокариотам. Цианобактерии - самая обширная, наиболее богатая формами и самая
распространенная группа фотосинтезирующих прокариот. Благодаря способности
расти в экстремальных условиях и фиксировать молекулярный азот они приобрели
большое значение в сложном «хозяйстве» природы. Некоторые
цианобактерии подвижны. Однако передвижение их никогда не осуществляется с
помощью жгутиков, а только путем скольжения по твердой поверхности.
Морфология
и систематика. Цианобактерии включают одноклеточные и многоклеточные
формы, и по морфологическим признакам их можно подразделить на пять групп (рис.
3.19). 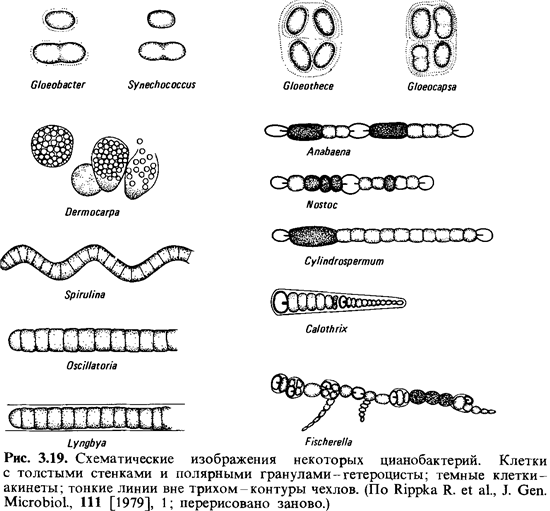
Группа 1: хроококковые
цианобактерии. Это одноклеточные палочки и кокки. Клетки существуют по
отдельности или в виде агрегатов (колоний), в которых они объединены капсулами
или слизью. Размножение клеток происходит исключительно бинарным делением или
почкованием. К этой группе относят Synechococcus (прежнее
название - Anacystis nidulans), Gloeothece и Gloeobacter violaceus.
Группа 2: плеврокапсовые
цианобактерии. Это тоже одноклеточные формы, но только такие, которые
могут размножаться и множественным делением. При этом внутри делящейся клетки
появляется много маленьких клеток, так называемых беоцитов. В качестве примеров
можно упомянуть роды Pleurocapsa, Dermocapsa и Myxosarcina.
Последующие
три группы характеризуются нитчатыми объединениями клеток; эти бактерии
образуют трихомы, т.е. цепочки из клеток. Рост осуществляется интеркалярно -
путем деления клеток внутри трихомы. Трихомы способны двигаться путем
скольжения. В ряде случаев при размножении происходит «разлом» трихомы и
образование гормогониев. По этой причине нитчатые формы иногда называют гормогониевыми
сине-зелеными водорослями. В настоящее время различают три группы
цианобактерий, образующих трихомы.
Группа 3: нитчатые
цианобактерий без гетероцист. Трихомы состоят только из вегетативных
клеток. Типичными для группы являются Oscillatoria (известная
«осциллирующая водоросль»), Spirulina, Lyngbya, Phormidium и Plectonema.
Группа 4: нитчатые
цианобактерий с гетероцистами. В трихомах,
растущих в отсутствие связанного азота, происходит дифференциация клеток с
образованием гетероцист. У некоторых форм встречаются также акинеты. К этой
группе относятся роды Anahaena, Nostoc и Calothrix.
Группа 5: это
тоже нитчатые цианобактерий с гетероцистами. Представители этой
группы отличаются от предыдущего типа делением клеток более чем в одной
плоскости. Наиболее известен род Fischerella.
Экология. Цианобактерий
распространены в озерах и других водоемах, в почве и на рисовых полях. Невооруженным
глазом их можно увидеть в виде темно-синей или черной пленки на скалах, в зоне
прибоя по берегам пресноводных озер и на морской литорали. Черные («чернильные»)
полосы на известняковых скалах, характерные для мест, где стекает вода, обязаны
своим происхождением хроококковым цианобактериям. В эвтрофных озерах часто
бывают вспышки массового размножения сине-зеленых (Anabaena) или
красных (Oscillatoria rubescens) цианобактерий, так называемое
цветение воды. Благодаря своей способности фиксировать молекулярный азот многие
формы цианобактерий первыми заселяют места, бедные питательными веществами, на
пример морской песок или скалы в пустыне, где в узких щелях они на ходят защиту
и влагу. Они не боятся и иных экстремальных условий. Некоторые одноклеточные
цианобактерии (Synechococcus lividus) на столько устойчивы к
действию кислот и термофильны, что способны расти в кислых горячих источниках
(рН 4,0, 70°С). Другие виды растут как симбионты: Nostoc в составе
лишайника Peltigera, а также в корнях Cycas и Gunnera; Anabaena
azollae – в полостях
листьев тропического водного папоротника Azolla.
Клетка, ее
строение и субклеточные компоненты. В отношении
тонкой структуры клеток цианобактерии во многом сходны с грам-отрицательными
бактериями (рис. 2.4). Протопласт окружен клеточной стенкой, в которой поверх
пептидогликанового слоя имеются еще «наружная мембрана» и липополисахаридный
слой. У многих форм имеет место выделение экзополисахаридов, которые либо
переходят в раствор в виде слизи, либо образуют капсулы вокруг
клеток и чехлы вокруг трихом. 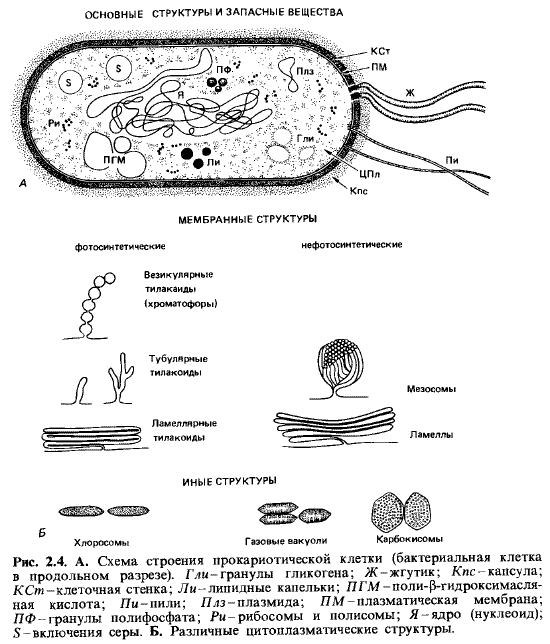
Фотосинтетический
аппарат представлен тилакоидами, которые либо расположены параллельно
плазматической мембране, либо сильно извиты и помещаются в периферических
участках цитоплазмы (рис. 3.20). Тилакоидная мембрана содержит хлорофилл а, Р-каротин
и оксокаротиноиды, такие как миксоксантофилл, эхиненон и зеаксантин, а также
компоненты цепи фотосинтетического переноса электронов. Отличительной
особенностью цианобактерии (и красных водорослей) является наличие фикобилисом.
Это дисковидные образования, прилегающие снаружи к тилакоидам. Они состоят из
фикобилипротеинов, чаще всего - фикоцианина (75%), аллофикоцианина (12%) и
фикоэритрина, а также бесцветных полипептидов, которые все вместе составляют
12%. Фикобилипротеины в свою очередь состоят из белка и простетических групп -
фикоцианобилина или фикоэритробилина. По своей функции это светособирающие
пигменты цианобактерии; поглощенная ими энергия передается главным образом
фотосистеме II. Хлорофилл а снабжает энергией только
фотосистему I. Фикобилисомы могут составлять до половины всего белка клетки. 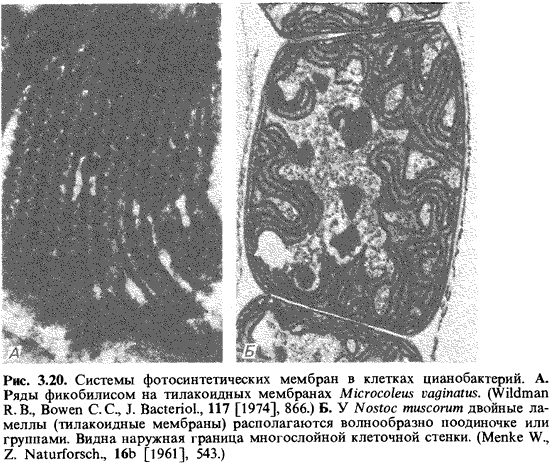
Фикобилины во
многом сходны с желчными пигментами. Их синтез начинается с образования
порфирина; при размыкании кольца углерод из метинового мостика выделяется в
виде СО. Синтез фикобилинов - один из немногих известных процессов, при которых
образуется окись углерода.
Только у
одной цианобактерий – Gloeobacter violaceus - нет тилакоидов и
фикобилисом; хлорофилл а находится в плазматической мембране, а
фикобилипротеины располагаются непосредственно у внутренней поверхности
мембраны сплошным слоем.
У многих
цианобактерий соотношение синего и красного пигментов зависит от спектрального
состава света. При зеленом и синем свете образуется преимущественно
фикоэритрин, при красном - фикоцианин. Такая комплементарная «хроматическая
адаптация» помогает эффективно поглощать свет в различных местообитаниях (под
пологом листвы или в голубом свете в глубине водоема).
Клеточные
включения. По-видимому, все цианобактерий способны к накоплению
полисахарида (в виде гликогеновых гранул) и гранул полифосфата.
Поли-р-гидроксимасляную кислоту накапливают лишь немногие виды.
Запасным
веществом, встречающимся только у цианобактерий, являются гранулы
циаиофицина. Они связывают вещества, окрашивающие белки, и это выдает
их полипептидную природу. У полипептида, построенного из остатков аспарагиновой
кислоты, все свободные карбоксильные группы последней связаны с аргинином;
таким образом, молекула цианофицина состоит из аспартата и аргинина в
соотношении 1:1. Этот полимер, очевидно, служит запасным источником азота; при
недостатке азота количество полимера убывает, а при добавлении в среду
источника азота оно вновь возрастает. Цианофицин накапливается главным образом
в гетероцистах. Он может также служить энергетическим резервом (хотя и
незначительным), так как аргинин и в анаэробных условиях расщепляется с
образованием орнитина и карбамоилфосфата, что в конечном счете приводит к
регенерации АТР.
Карбоксисомы
у цианобактерий очень широко распространены. Водные формы цианобактерий,
обитающие в стратифицированных озерах, а также вызывающие цветение воды, почти
все имеют газовые вакуоли.
Специализированные
клетки. У цианобактерий имеются сильно дифференцированные клетки,
которым нет аналогов ни в одной другой группе бактерий. Рассмотрим отдельные
типы таких клеток. 
Гетероцисты уже
при исследовании в световом микроскопе обратили на себя внимание своими
толстыми клеточными стенками, слабой пигментацией и сильно преломляющими свет
полярными гранулами. Электронная микроскопия дала возможность изучить их тонкую
структуру (рис. 3.21). Что касается полярных гранул, то они оказались цианофициновыми
гранулами; плотные же слои, расположенные поверх грам-отрицательной клеточной
стенки, состоят из полисахаридов, в которых глюкоза, галактоза, манноза и
ксилоза соединены между собой р-1,3-гликозидными связями. Гетероцисты устойчивы
к действию лизоцима. С соседними клетками трихомы гетероцисты связаны порами,
своего рода плазмодесмами. По своей функции гетероцисты - это места
фиксации азота (N2) в аэробных условиях. Они образуются у
нитчатых цианобактерий при недостатке связанного азота (NH2, NO2).
Одновременно с морфологической дифференциацией происходят биохимические изменения.
В гетероцистах синтезируется нитрогеназа, а фикобилипротеины распадаются, тогда
как хлорофилл а сохраняется. Таким образом, зрелые гетероцисты не
содержат ни фикобилипротеинов, ни функционирующей фотосистемы II и поэтому не
могут выделять О2. Они имеют только фотосистему I, дающую им
возможность осуществлять так называемое циклическое фотофосфорилирование и
регенерацию АТР. В гетероцистах, таким образом, создаются все условия,
необходимые для функционирования нитрогеназы, которая чувствительна к кислороду.
Соединениями углерода их снабжают вегетативные клетки, а связанный азот
гетероцисты отдают преимущественно в форме глутамина.
Акинеты представляют
собой покоящиеся клетки. Они выделяются своими размерами, сильной пигментацией
и толстой клеточной стенкой. Подобно гетероцистам, это морфологически
дифференцированные клетки; они могут быть расположены в середине трихомы (у Апаbаепа) или на ее
конце (у Cylindrospermum), внутри или на поверхности (рис.
3.19).
Гормогонии
- короткие отрезки, получающиеся при разрыве длинных трихом. Они служат для
размножения. При образовании гормогония осцилляториям приходится жертвовать по
меньшей мере одной клеткой трихомы, так как клетки связаны общим
пептидогликановым слоем, который не может быть разорван на границе клеток. В
этом случае говорят о трансцеллюлярном разрыве.
Беоциты («мелкие
клетки») - это репродуктивные клетки плеврокапсовых цианобактерий. Они
образуются в результате множественного бинарного деления материнской клетки - очень
крупной клетки с толстой волокнистой экзополисахаридной оболочкой. Примером
может служить Dermocarpa, у которой быстро, одно за другим,
происходят бинарные деления и из одной материнской клетки получается от 4 до
1000 беоцитов.
Скользящее
движение. Многие цианобактерий подвижны. Таковы, например, формы,
образующие трихомы, все гормогонии и многие беоциты. Они могут передвигаться
только путем скольжения, а для этого необходим твердый субстрат. Движение
сопровождается вращением трихомы вокруг продольной оси; при этом кажется, что
нити начинают колебаться, осциллировать (отсюда название Oscillatoria). Например,
у Oscillatoria princeps точка на поверхности трихомы
(поставленная тушью) в процессе продвижения описывает винтовую линию крутизной
в 60°. Исследование тонкой структуры клеточной стенки показало, что в ней
имеются фибриллы, проходящие под таким же углом крутизны. Это позволяет
предполагать, что движение основано на торзионных волнах, создаваемых
фибриллами. Направление движения может меняться на обратное. Многие цианобактерий
способны к фототаксису и концентрируются в области с подходящей для них
освещенностью.
Фиксация
азота. Связывать N2 способны все цианобактерий, образующие
гетероцисты. Гетероцисты - это клетки, морфологически и физиологически
специализированные для фиксации азота (см. выше). Неожиданным было открытие,
что генетическая информация (гены nif), необходимая для образования
нитрогеназы, имеется даже в клетках плеврокапсовых цианобактерий, хотя среди
них нет нитчатых форм, образующих гетероцисты (группа Oscillatoria). В
обычных условиях роста на свету фиксации N2 у них, правда, не
происходит, так как фермент нитрогеназа очень чувствителен к кислороду. Однако
с помощью особого приема удалось все - таки вызвать синтез нитрогеназы и обнаружить
этот фермент. Если клетки инкубировать в отсутствие связанного азота в
анаэробных условиях на свету и, добавив в среду дихлорфенил-метилмочевину,
подавить функцию фотосистемы II (так что выделение О2 прекратится),
то будет синтезироваться нитрогеназа. Таким способом удалось доказать, что
более половины изученных штаммов способно к образованию нитрогеназы.
Среди
одноклеточных хроококковых цианобактерий некоторые штаммы также способны
связывать N2. Каким образом у этих бактерий нитрогеназа защищена от
кислорода, выделяемого при фотосинтезе, пока неизвестно.
Анаэробный
метаболизм. Многие цианобактерий живут в слоях воды, содержащих в
значительных концентрациях (примерно 5 мМ/л) сероводород. Исследования,
проведенные на Oscillatoria limnetica, показали, что фотосистема II в
присутствии сероводорода выключается, и начинает протекать иной, бескислородный
(аноксигенный) фотосинтез по схеме
С02
+ 2H2S -> <СН20> +
Н20 + 2S
известной для
анаэробных пурпурных серобактерий. Но способность многих цианобактерий выживать
и в анаэробных условиях, вероятно, имеет значение лишь в немногих
местообитаниях.
Облигатная
фотоавтотрофия. По-видимому, многие цианобактерий - облигатные
фотоавтотрофы; они могут расти только на свету. Лишь некоторые штаммы могут
существовать и как хемоорганотрофы, окисляя сахара в темноте. В таких условиях
скорость их роста всегда намного меньше, чем в фотоавтотрофных условиях.
Прохлорон (Prochloron). Недавно
был найден новый фотосинтезирующий организм, соединяющий в себе ряд признаков
прокариот и зеленых водорослей. С одной стороны, у него типичная
прокариотическая клетка (грам-отрицательная клеточная стенка с пептидогликаном,
нет органелл и истинного ядра, размеры генома 3,6-109 Да); с
другой стороны, наряду с хлорофиллом а он имеет и хлорофилл b, свойственный
только зеленым водорослям и высшим растениям. От цианобактерий он отличается
также отсутствием фикобилипротеинов, цианофицина и поли-Р-гидроксимасляной
кислоты. Эта переходная форма получила название Prochloron, и
ее относят к самостоятельной группе протохлорофитов. Prochloron растет
как эктосимбионт на различных асцидиях.
| 



