Спонтанные мутации
В популяции бактерий без всякого экспериментального вмешательства регулярно возникают мутации; такие мутации называют спонтанными мутациями, а клетки, в которых они возникли,-спонтанными мутантами. Мутагенное действие аналогов оснований ДНК (см. ниже) указывает на возможные причины спонтанных мутаций: вероятно, речь идет о случайных ошибках при включении нуклеотидов во время репликации ДНК-ошибках, вызванных таутомерным перемещением электронов в основании. Тимин, например, обычно находится в оксо-форме, в кото рой он образует водородные связи с аденином. Но если тимин во время спаривания оснований при репликации ДНК переходит в енольную форму, то он спаривается с гуанином. В результате в новой молекуле ДНК на том месте, где раньше находилась пара А-Т, появляется пара G-C.
Доля мутантов в популяции и частота мутирования. Численная доля мутантов в клеточной -популяции для разных признаков различна и варьирует в пределах от 10 ~4 до 10" и. Она зависит от частоты возникновения мутаций, условий среды, возраста клеточной суспензии и других факторов. Вероятность возникновения определенных мутаций в расчете на одну клетку и на одну генерацию называют частотой мути рования. При высоких скоростях роста она постоянна, и ее обычно определяют для клеток в экспоненциальной фазе роста при опти мальных условиях среды. Частота спонтанных мутаций для определен ного гена составляет величину порядка 10" 5, а для определенной пары нуклеотидов 10 ~ 8.
«Молчащие» мутации. Если под мутацией в традиционном смысле понимают внезапное изменение признака, т.е. изменение генотипа, про являющееся в фенотипе, то на молекулярном уровне любое стабильное наследуемое изменение ДНК рассматривают как мутацию. Однако вви ду вырожденности генетического кода понятно, что не всякая мутация такого рода будет проявляться в фенотипе. Во многих триплетах изме нение третьего основания остается без последствий («молчащие» мута ции). Даже замена первого или второго основания триплета не всегда приводит к серьезным последствиям. Хотя структуры высшего порядка (третичная и четвертичная) определяются первичной структурой белка (т.е. последовательностью аминокислот), разные аминокислоты играют в этой структуре не одинаково важную роль. Например, мутация AUC->GUC ведет к замене изолейцина валином, т.е. к замене одной липофильной группы на другую. Однако мутация CUU->CCU приве дет к замене лейцина пролином, и последствием такой замены будет от клонение от нормальной пространственной конфигурации полипептид ной цепи, что может сильно изменить структуру высшего порядка. Из этого понятно, что различные мутации в одном и том же структурном гене определенного фермента могут по-разному сказываться на его ак тивности: возможны любые изменения-от едва заметного снижения ка талитического действия до полной инактивации.
Обратные мутации и реверсии. Из сказанного выше становится ясно, что у мутанта может произойтиобратная мутация, в результате кото рой восстановятся свойства дикого типа. Об истинной обратной мутации говорят лишь в тех случаях, когда вторая мутация точно восстана вливает исходный генотип, т.е. когда измененный при первой мутации триплет будет вновь кодировать ту же аминокислоту, что и раньше. Ее ли же дело сводится к восстановлению исходного фенотипа (например, к возобновлению синтеза нормально функционирующего фермента), то говорят о реверсии или супрессорной мутации и соответственно оревер-тантах. Супрессорные мутации могут происходить как в исходном гене, так и в каких-либо других участках хромосомы (интрагенные и экстрагенные супрессорные мутации).
Индуцированные мутации
Обрабатывая клетки мутагенными (вызывающими мутации) вещества ми, можно повысить частоту мутаций. В этом случае говорят об индук ции мутаций, а полученные при этом клетки называют индуцированны ми мутантами. Мутагенами могут быть химические, физические или биологические агенты. Механизм их действия будет пояснен на ряде примеров.
В отношении генетической структуры различают три класса мутан тов со следующими дефектами: 1) одна пара оснований заменена дру гой, например вместо AT может быть GC или наоборот; 2) включена дополнительная пара оснований в нуклеотидную последовательность или утрачена одна из существовавших пар; 3) группа оснований или да же генов может быть утрачена (делеция), перемещена в пределах хромосомы (транспозиция) или «разорвана» путем вставки посторонней ДНК (инсерция).
Для мутаций класса 1, называемых также точечными мутациями, характерна высокая частота реверсии. В случае мутаций класса 2, к ко торым относятся также мутации со сдвигом рамки (см. рис. 15.4), ревер-танты редки, а после мутаций класса 3 (за некоторыми исключениями) ревертанты не появляются.
В последующих разделах мы остановимся на некоторых механизмах мутагенеза.
Включение аналогов оснований. Аналоги оснований - это антиметабо литы. Некоторые аналоги настолько сходны с нормальными пиримиди-новыми и пуриновыми основаниями, что поглощаются клетками и включаются в ДНК. Здесь они в значительной степени выполняют функцию нормальных оснований, но в отличие от них обнаруживают большую тенденцию связывать «ложного» (неподходящего) партнера при репликации ДНК. Для вызывания мутаций часто используются бромурацил и 2-аминопурин. Бромурацил представляет собой соедине ние, аналогичное по структуре тимину, которое включается вместо него в цепь ДНК как партнер аденина (рис. 15.3). Бромурацил таутомери-зуется в енольную форму чаще, чем тимин. При репликации цепи, со держащей бромурацил, он в енольной форме спаривается как цитозин, т.е. вызывает включение гуанина вместо аденина. Таким образом, в не которых случаях пара оснований AT заменяется на CG. 2-Аминопурин включается в ДНК вместо аденина и действует подобным же образом. Этот вид изменений - замену одного пурина другим пурином (A->G)

или одного примидина другим пиримидином (С-> Т) - называют тран-зицией.
Химическое изменение оснований. Некоторые мутагенные вещества действуют путем химического изменения содержащихся в ДНК основа ний, что приводит к ошибкам репликации. Вполне понятное изменение вызывает нитрит. Азотистая кислота дезаминирует аденин, гуанин или цитозин без разрыва или каких-либо других изменений полинуклеотид-ной цепи. В результате замещения аминогруппы гидроксильной группой аденин превращается в гипоксантин и спаривается с цитозином вместо тимина, что приводит к мутации AT -► GC. Если цитозин дезаминирует-ся в урацил. то он спаривается с аденином вместо гуанина, и это ведет к мутации GC -»AT. Будучи превращен в ксантин, гуанин по-прежнему спаривается с цитозином, т. е. дезаминироваиие G не вызывает мутации. Гидроксиламин вступает в реакцию главным образом с цитозином и из меняет его так, что тот спаривается с аденином; значит, он тоже вызы вает мутации CG->TA.
Алкилирующие агенты. Этил- и метилметансульфонат, диметил-и диэтилсульфат, этиленимин, азотистый или серный иприт, а также N-метил-1ЧГ'-нитро-]ЧГ-нитрозогуанидин принадлежат к наиболее эффек тивным мутагенам. Например, этилметансульфонат этилирует преиму щественно атом N гуанина. Образовавшийся 7-алкилгуанин отщепляет ся от цепи, в результате чего в ней образуется «пропуск». При очередной репликации на этом месте часто оказывается «ошибочное» основание.
Включение или утрата отдельных пар оснований. Профлавин и другие акридиновые красители действуют по-иному. Вероятно, молекула акридина внедряется между соседними основаниями цепи ДНК и увели чивает расстояние между ними (интеркаляция). Такое пространственное изменение при репликации ДНК может вызывать ошибки двух типов-

утрату нуклеотида или включение дополнительной пары нуклеотидов. Мутации этого типа приводят к очень серьезным последствиям, так как при этом нарушается порядок считывания информации при синтезе бел ка: начиная с места утраты или включения нуклеотида, информация считывается в «неправильных» триплетах (мутация «со сдвигом рамки», рис. 15.4).
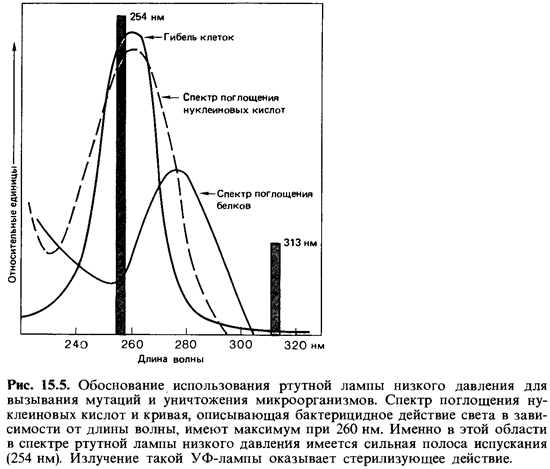
Ультрафиолетовые лучи и ионизирующее излучение. УФ-свет, рентге новские лучи и другие виды ионизирующего излучения оказывают на микроорганизмы как подавляющее жизнедеятельность (летальное), так и мутагенное воздействие. Их специфическое действие еще мало изуче но. Исходя из совпадения кривой поглощения нуклеиновых кислот и кривой подавления жизнедеятельности клеток при облучении в зави симости от длины волны, а также частоты мутаций в популяции, можно сделать вывод о том, что УФ-лучи действуют в основном на нуклеи новые кислоты. Наиболее эффективны лучи ближней УФ-области с дли ной волны около 260 нм (рис. 15.5). Побочные повреждения при этом незначительны. Поражаются главным образом пиримидиновые основа ния. Например, два соседних тиминовых основания в ДНК могут ока заться ковалентно связанными. Наличие таких димеров тимина служит затем источником ошибок при репликации (рис. 15.6).
Репарация ДНК. На подвергнутых УФ-облучению бактериях было показано, что повреждения ДНК частично обратимы. Если облучить бактериальную сус пензию большой дозой ультрафиолета, то значительная доля клеток подверг нется летальному повреждению (в них возникнут летальные мутации); при нача той сразу же инкубации в темноте лишь немногие клетки образуют колонии. Но если непосредственно после УФ-облучения воздействовать на клетки светом бо-
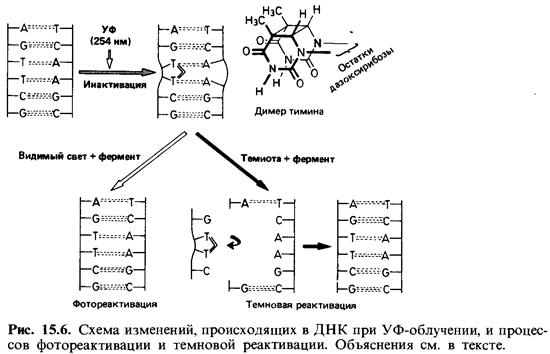
лее длинноволновой области (320-550 нм), то доля выживших клеток возрастет в несколько десятков раз. В такой фотореактивации участвует фермент, активи руемый светом и восстанавливающий нормальную структуру ДНК путем рас щепления образовавшихся димеров тимина. Есть и другой репаративный меха низм, не требующий участия света. При этой темповой реактивации дефектные участки цепи ДНК вырезаются и заменяются новыми нуклеотидами. Степень эффективности такой репарации лучевых повреждений у разных штаммов бакте рий неодинакова. Устойчивость к облучению, характерная для некоторых бакте рий (например, Micrococcus radiodurans), обусловлена высокоэффективным меха низмом репарации.
Мутации, вызываемые транспозонами. В генетике бактерий все боль шее значение приобретает метод получения мутаций с помощью транс-позонов. Транспозоны (Тп) представляют собой короткие двойные цепи ДНК, которые состоят из более чем 2000 пар оснований и обычно обус ловливают устойчивость к одному антибиотику, в исключительных слу чаях-к нескольким. Транспозоны способны «перепрыгивать» из одного участка генома в другой, в частности из бактериальной хромосомы в плазмиду и обратно; таким образом, они могут включаться в раз личные участки генома (см. разд. 15.3.1). В случае внедрения транспозо-на в какой-либо структурный ген хромосомы нуклеотидная последова тельность этого гена будет нарушена и генетическая информация не сможет транслироваться в функционально полноценный полипептид. Возникнет инсерционный мутант.
Поскольку транспозоны не способны к автономной репликации, для переноса их из одной бактериальной клетки в другую необходим так называемый вектор (переносчик). Векторами могут служить плазмиды или бактериофаги. Следует упомянуть, что колифаг мю («фаг-мута-тор»), подобно транспозону, обладает способностью внедряться в раз личные участки бактериальной хромосомы и вызывать мутации. По этой причине фаг мю был назван «гигантским транспозоном», и его ис пользуют в повседневной практике получения мутантов Е. coli.
Проявление признаков. Уже возможность фотореактивации после УФ-облучения указывает на то, что первичный эффект при воздействии мутагенного фактора не обязательно ведет к истинной мутации. Включение бромурацила в цепь ДНК или димеризация тимина представляет собой лишь премутацию;димеризация тимина - процесс обратимый, и в случае фотореактивации дело не доходит до возникновения мутанта. Только при последующей редупликации премутировавшей цепи ДНК первичное повреждение становится стабильным и в дальнейшем пере дается потомству как новый элемент генотипа. Такая закрепившаяся мутация может исчезнуть только в результате обратной мутации. Про явление мутации в фенотипе связано с рядом последовательных процес сов, которые требуют определенного времени или нескольких кле точных делений. Новый фенотип проявится лишь тогда, когда измененный ген начнет функционировать. Этапы, необходимые для ре ализации нового фенотипа, различны для разных клеток и разных типов мутаций.
Запаздывающее проявление мутаций. Если в гаплоидной клетке про изойдет реверсия, превращающая ауксотрофную мутантную клетку в прототрофную, то такая обратная мутация сразу проявится в феноти пе. Восстановление способности вырабатывать определенный фермент можно в надлежащих условиях тотчас же распознать. Иначе обстоит де ло с мутациями, приводящими, наоборот, к ауксотрофному состоянию, например к утрате способности синтезировать определенную аминокис лоту. Такие мутации удается распознать лишь по прошествии периода, включающего несколько клеточных генераций. Запаздывающее проявле ние объясняется в данном случае тем, что, хотя мутация и делает невоз можным синтез необходимого фермента, еще продолжает какое-то вре мя действовать фермент, синтезированный ранее. Новый признак проявится лишь тогда, когда в результате клеточных делений произой дет достаточное «разбавление» этого фермента. С запаздывающим из менением фенотипа приходится также считаться при выявлении фаго-устойчивых бактерий. Если фагочувствительные бактерии приобретают устойчивость в результате мутации, ведущей к утрате способности син тезировать особое рецепторное вещество, то эта устойчивость выявится лишь тогда, когда в результате ряда клеточных делений это вещество будет в достаточной мере разбавлено.
У многоядерных (ценоцитных) клеток тоже следует различать мута ции, приводящие к приобретению и к утрате какой-либо функции. Мно гие бактерии имеют по нескольку ядер (хромосом). Клетки Escherichia coliпри быстром росте на средах, богатых питательными веществами, содержат в среднем по четыре хромосомы. При мутации с приобрете нием функции доминирует мутантная хромосома: она сразу же вызывает синтез нового фермента, и мутация немедленно проявляется в фе нотипе. Если же в многоядерной клетке произошла мутация с утратой функции, она оказывается рецессивной. При многократном делении кле ток ядра распределяются по разным дочерним клеткам (сегрегация ядер, рис. 15.7). Дефект может проявиться лишь в той клетке, у которой все
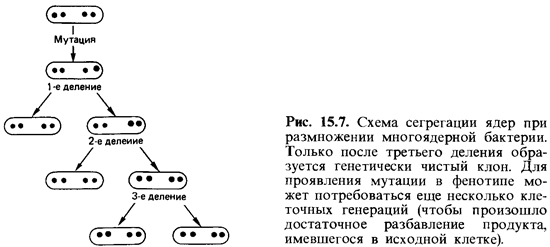
ядра содержат мутировавший ген. Потомство такой клетки предста вляет собой генетически чистый клон. Таким образом, при мутациях с утратой функции в случае многоядерных клеток следует учитывать как сегрегацию ядер, так и проявление мутации в фенотипе.




