Метан
образуется при анаэробном разложении органических веществ. Его запасы весьма
значительны; примерные подсчеты показывают, что 1-1,5% углерода, поступающего в
результате минерализации органиче ских веществ в виде С02 в
атмосферу, сначала попадает туда в виде ме тана и лишь затем превращается под
воздействием гидроксильных ра дикалов (ОН') в СО, а потом в С02. К
экосистемам, в которых образуется метан, относятся большие площади, занятые
тундрой и бо лотами (отсюда другое название метана-болотный газ); рисовые поля;
осадки на дне прудов и озер; лиманы, марши и эстуарии; отстойники очистных
сооружений и, наконец, желудки (рубцы) более чем 109 жвачных
животных. В анаэробных условиях органические вещества сна чала через ряд
промежуточных этапов сбраживаются до ацетата, С02 и Н2.
Эти продукты метаболизма первичных и вторичных деструкторов используются
метанобразующими (метаногенными) бактериями.
Систематическое
положение метанобразуюших бактерий. По морфо логии эти бактерии можно
подразделить на палочковидные (Methanobacterium), кокковидные (Methanococcus), сарциноподобные(Methanosarcina) и
спирилловидные (Methanospirillum) (табл. 9.2). 
Метанобразующие
бактерии составляют обособленную группу ми кроорганизмов. Они отличаются от
других бактерий не только типом метаболизма, но и некоторыми признаками,
касающимися состава кле точных структур. У них
нет типичного пептидогликанового остова:
Methanococcus обладает
лишь белковой стенкой; у Methanospirillum имеется полипептидный
чехол; клеточная стенка Methanosarcina barkeri построена
из полисахарида, в состав которого входят уроновые кис лоты, нейтральные сахара
и аминосахара. Пенициллин не подавляет ро ста метанобразующих бактерий.
Цитоплазматическая
мембрана содержит липиды, состоящие из эфи-ров глицерола и изопреноидных
углеводородов. Рибосомы по своей величине сходны с рибосомами
эубактерий (70S), однако после довательность оснований в рибосомных РНК,
особенно в 16S-pPHK, су щественно иная, чем у эубактерий. В отношении
последовательности 16S-pPHK метанобразующие бактерии значительно сильнее
отличают ся, например, от Е. coli, чем цианобактерии.
Кроме того, механизм трансляции нечувствителен к антибиотикам, подавляющим
синтез белка у эубактерий. На основании этих, а также ряда других отличительных
признаков метанобразующие бактерии относят к архебактериям.
Физиология. Метанобразующие
бактерии-строгие анаэробы: кисло род воздуха убивает их. У них нет ни каталазы,
ни супероксиддисму-тазы. Именно из-за высокой чувствительности этих бактерий к
кислоро ду наши сведения об их физиологии, биохимии и экологии пока
сравнительно скудны. Только после разработки специальных методов (например,
метода Хангейта) появилась возможность пересевать и выде лять метанобразующие
бактерии без доступа кислорода.
Большинство
до сих пор выделенных видов в чистой культуре спо собно использовать в качестве
донора водорода Н2, а некоторые из них-также формиат, метанол,
ацетат или метиламин. В ряде анаэ робных экосистем основным субстратом для
образования метана слу жит ацетат. Таким образом, круг субстратов очень узок.
Метанобразующие
бактерии составляют последнее звено анаэробной пищевой цепи , в начале которой находятся полисахариды (цел люлоза, крахмал), белки и жиры.
В этой цепи участвуют также 1) бакте рии, сбраживающие целлюлозу до сукцината,
пропионата, бутирата, лактата, ацетата, спиртов, С02 и Н2;
2) ацетогенные бактерии, сбражи вающие эти первичные продукты брожения до
ацетата, формиата, С02и Н2 (эти вещества в свою
очередь служат субстратами для ме танобразующих бактерий).
Метанобразующие
бактерии, очевидно, находятся в тесном взаимо действии с бактериями,
выделяющими водород (рис. 9.6). В микросреде обитания таких бактерий
газообразный водород в свободном состоянии практически не встречается. Водород,
выделяемый бактериями и раство ренный в среде, сразу же поглощается
метанобразующими видами. Из вестно, что высокое парциальное давление Н2 подавляет
метаболизм и рост многих бактерий, образующих водород. Это означает, что не
только метаногенные бактерии зависят от продуцентов Н2, но и те в
свою очередь зависят от поглощающих Н2метанообразователей. Та ким
образом, имеет место ассоциация типа мутуалистического сим биоза. 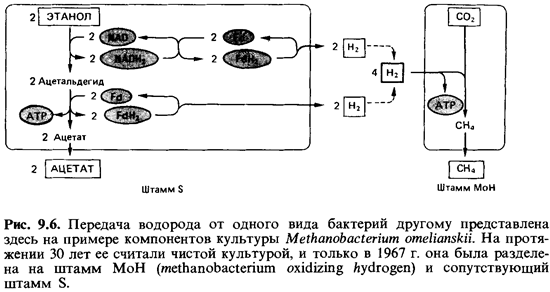
Метанобразующие
бактерии способны активировать водород и осу ществлять его окисление,
сопряженное с восстановлением С02. По скольку клеточные вещества
ряда видов могут синтезироваться из С02 как единственного
источника углерода, можно рассматривать способ су ществования этих бактерий как
хемоавтотрофный. Для получения энер гии С02 используется в
качестве акцептора водорода, что ведет к обра зованию метана:
Таким
образом, метанообразование можно по аналогии с другими видами дыхания называть
карбонатным дыханием. Поэтому метан образующие виды характеризуют как
анаэробные автотрофные бактерии, окисляющие водород:
4Н2
+ С02 -» СН4 + 2Н20;
AG0 =
- 131 кДж/моль ( - 31,3 ккал/моль)
Некоторые
метанобразующие бактерии могут превращать в метан и окись углерода. При этом в
качестве промежуточных продуктов обра зуются СО2 и Н2:
4С0 +
4Н20--------- ►4С02+4Н2
С02 +
4Н2 -------- * СН4 +
2Н20
4С0 +
2Н20--------- ► СН4+ЗС02
Биохимия образования
метана и получения энергии. В биохимическом превращении Н2 и
С02 в метан или ацетата в метан и С02 участвует
ряд коферментов и простетических групп, которые до сих пор были найдены только
у метанобразующих бактерий: производное деазарибо-флавина F420,
метаноптерин, метанофуран, никельтетрапиррольный фактор F430 и кофермент
М (меркаптоэтансульфонат).
Пока мало что
известно о ферментах, участвующих в отдельных реак циях. Вопрос о регенерации АТР тоже
окончательно еще не выяснен. С точки зрения термодинамики только последняя
реакция, приводящая к образованию метана, может сопровождаться синтезом АТР.
Экспери менты на Methanosarcina barkeri-бактерии, способной
также осущест влять реакцию СН3ОН + Н2 -» СН4 +
Н20,-дали следующие резуль таты. Добавление обоих субстратов к
бактериальной суспензии приводило к выведению протонов, образованию АТР и
синтезу метана. При участии метилтрансферазы метанол может прямо превращаться в
метилкофермент М, при восстановлении которого образуется метан:
СН3-S-СоМ
+ Н2 - СН4 +
HS = СоМ
Восстанавливающий
фермент - метил-СоМ-метилредуктаза-предста вляет собой мультиферментный
комплекс, который содержит, в частно сти, белковые факторы F420, F430 и
гидрогеназу. Вероятно, реакция всегда сопровождается выведением из клетки
протонов, и создающийся в результате этого протонный потенциал доставляет
энергию для реге нерации АТР. Из этих результатов можно заключить, что вообще
метанобразующие бактерии синтезируют АТР не путем фосфорилиро-вания на уровне
субстрата, а путем окислительного фосфорилирования в анаэробных условиях
(«анаэробное дыхание»).
Биохимия
ассимиляции С02. Автотрофная фиксация С02 у
метан-образующих бактерий [так же как и у сульфатредуцирующих и
ацетогенных анаэробов, использующих водород] происходит без участия
реакций рибулозобисфосфатного цикла. Путь синтеза кле точных веществ из С02 идет
через синтез ацетил-СоА и пирувата. Этапы этого пути были установлены с помощью
радиоактивных соеди нений, а также в результате исследования ферментов Methanobacterium thermoautotrophicum. Механизмы
этих превращений в настоящее время интенсивно изучаются.
СО 2 восстанавливается
до метанола (в связанной форме). Вторая моле кула С02 восстанавливается
с помощью СО-дегидрогеназы до СО. Вос становительные эквиваленты генерируются
путем активации Н2гидро-геназами и переносятся ферментами,
реагирующими с фактором F420 или с NADP. Карбонилирование
метил-Х ведет к образованию аце-тил-Х, а в результате восстановительного
карбоксилирования ацетил-СоА с помощью пируватсинтазы получается пируват, из
которого из вестными путями синтезируются клеточные вещества.
Практическое
значение метанобразующих бактерий. Отстойники, в ко торых происходит
анаэробный распад органических веществ сточных вод населенных пунктов,
относятся к обычному оснащению коммунальных водоочистных сооружений. В
индустриальных странах гниение ила служит в первую очередь стабилизации
первичного ила, а также ак тивного ила, образующегося при аэробной очистке
сточных вод. Метан, выделяющийся при гниении ила, частично используется
определенными микроорганизмами, а частично его применяют в качестве топлива. В
сельском хозяйстве применяют биогаз-ферментеры, а также ямы для навоза, чтобы
сбраживать (с получением метана) экскременты жи вотных вместе с отбросами,
содержащими целлюлозу. Метод образова ния биогаза, основной частью которого
является метан, обладает двумя важными преимуществами: во-первых, сохраняется
содержащийся в эк скрементах азот, а также те ценные качества удобрения,
которыми обладает сероводородный ил; во-вторых, происходит образование био газа
(в основном метана), который может быть использован в качестве источника
энергии как в сельскохозяйственном производстве, так и в домашнем хозяйстве. | 



