| Пятница, 01.11.2024, 01:59 | Главная | Регистрация | Вход |
|
Статистика

Онлайн всего: 1 Гостей: 1 Пользователей: 0 |
Статьи
В разделе материалов: 204
Показано материалов: 41-50 |
Страницы: « 1 2 3 4 5 6 7 ... 20 21 » |
При внесении
бактерий в питательную среду они обычно растут до тех пор, пока содержание
какого-нибудь из необходимых им компонентов среды не достигнет минимума, после
чего рост прекращается. Если на протяжении этого времени не добавлять
питательных веществ и не удалять конечных продуктов обмена, то получим так
называемую периодическую культуру (популяцию клеток в ограниченном жизненном
пространстве). Рост в такой «закрытой системе» подчиняется закономерностям,
действительным не только для
одноклеточных, но и для многоклеточных
организмов. Периодическая культура ведет себя как многоклеточный организм с
генетически ограниченным ростом. |
Для изучения
метаболических процессов на протяжении цикла клеточного деления нужны такие
суспензии, в которых клетки делились бы одновременно (синхронно). Чтобы достичь
такого совпадения фаз цикла у разных клеток, прибегают к синхронизации
культуры. Синхронизировать деление в какой-либо популяции клеток можно с
помощью различных искусственных приемов, таких как изменение температуры, воздействие
света, ограничение количества питательных веществ или пропускание
микроорганизмов через специальный фильтр с целью по лучить клетки одного
размера. Клеточная суспензия, синхронизированная той или иной обработкой, после
нескольких одновременных делений постепенно переходит снова к асинхронному
делению, так что число клеток увеличивается в дальнейшем уже не ступенчато, а
непрерывно. |
Ряд
химических агентов замедляет или полностью тормозит рост микроорганизмов. Если
то или иное вещество подавляет рост бактерий, а после его удаления рост вновь
возобновляется, то говорят о бактериостатическом действии. Бактерицидные вещества
вызывают гибель клеток. Тот или другой эффект, однако, зависит от концентрации
действующего вещества. Кроме того, среди бактерий существуют формы, устойчивые
к общим клеточным и метаболическим ядам (таким, как сероводород, фенол или
окись углерода) и даже способные использовать их как источники энергии. Для
многих антимикробных агентов удалось в той или иной мере выяснить субклеточную
мишень и механизм действия. |
Уничтожение
микроорганизмов - один из необходимых элементов микробиологической работы и
основа консервирования пищевых продуктов; поэтому стоит остановиться на нем
подробнее. Освобождение какого-либо материала от живых микроорганизмов или их
покоящихся форм называют обеспложиванием или стерилизацией. От стерилизации
следует отличать частичное обеспложивание (пастеризацию), а также консервирование.
Если стерильная среда или микробная культура загрязняется случайно попавшими в
нее микроорганизмами, то говорят о контаминации, или загрязнении. Такие
понятия, как дезинфекция (уничтожение всех патогенных микроорганизмов), асептика
и антисептика, а также инфекция, употребляются главным
образом в гигиене, а не в микробиологии. |
Органические
материалы разлагаются микроорганизмами, если только против этого не приняты
специальные меры. Для сохранения органических веществ (для их консервирования)
можно использовать ряд средств. Наибольшее значение имеют методы,
обеспечивающие сохранность пищевых продуктов. Этими вопросами занимается
пищевая микробиология.
Пищевые
продукты становятся негодными к употреблению не только в результате их
разложения микроорганизмами (аэробного окисления или анаэробного гниения), но и
вследствие того, что на них поселяются бактерии и грибы, образующие токсины.
Важнейшими продуцентами токсинов, попадающими в пищевые продукты, являются Clostridium botulinum и
различные виды стафилококков. С. Botulinum выделяет
экзотоксин, чрезвычайно сильно действующий даже в малых количествах и
поражающий нервную систему. Стафилококки образуют энтеротоксин, вызывающий так
называемые пищевые отравления. Некоторые грибы образуют микотоксины, из которых
наиболее известен афлатоксин (продукт жизнедеятельности гриба Aspergillus flavus).
Методы
консервирования, применяемые для защиты пищевых продуктов от микроорганизмов,
весьма разнообразны. Используются как физические, так и химические методы. |
Рассматривая
круговорот углерода, мы уже противопоставляли друг другу два процесса: фотосинтез, при котором фиксируется СО2 и
выделяется О 2, и минерализацию органических
веществ, связанную с потреблением О2 и освобождением СО2.
Процессы эти противоположны. С точки зрения массообмена важнее всего переход
углерода из газообразного неорганического вещества в полутвердые и твердые
органические соединения и обращение этого процесса. Если же рассматривать оба
этих процесса с позиций энергообмена, т.е. преобразования энергии, то углерод
имеет меньшее значение, чем водород. |
Метаболизм
и метаболические пути. Как во время роста, так и в состоянии покоя
вегетативные клетки нуждаются в постоянном притоке энергии. Живая клетка
представляет собой высокоорганизованную материю. Энергия необходима не только
для создания такой организации, но и для ее поддержания. Эту энергию организм
получает в процессе обмена веществ, или метаболизма, т.е. путем регулируемых
превращений, которым различные вещества подвергаются внутри клеток. Источниками
энергии служат питательные вещества, поступающие из внешней среды. В клетках
эти вещества претерпевают ряд изменений в результате последовательных
ферментативных реакций, образующих этапы определенных метаболических путей.
Такие пути выполняют две главные функции: они, во-первых, поставляют материалы
- предшественники для построения клеточных компонентов и, во-вторых,
обеспечивают энергию для клеточных синтезов и других процессов,
требующих затраты энергии. |
Несколько
путей ведут от глюкозы к С3-соединениям и среди них к пирувату - одному
из важнейших промежуточных продуктов метаболизма. Чаще других используется путь
распада через образование фруктозе-1,6-дисфосфата; его называют фруктозодисфосфатным
путем, гликолитическим расщеплением, гликолизом или (по имени изучивших его
исследователей) путем Эмбдена-Мейергофа-Парнаса (рис. 7.3). 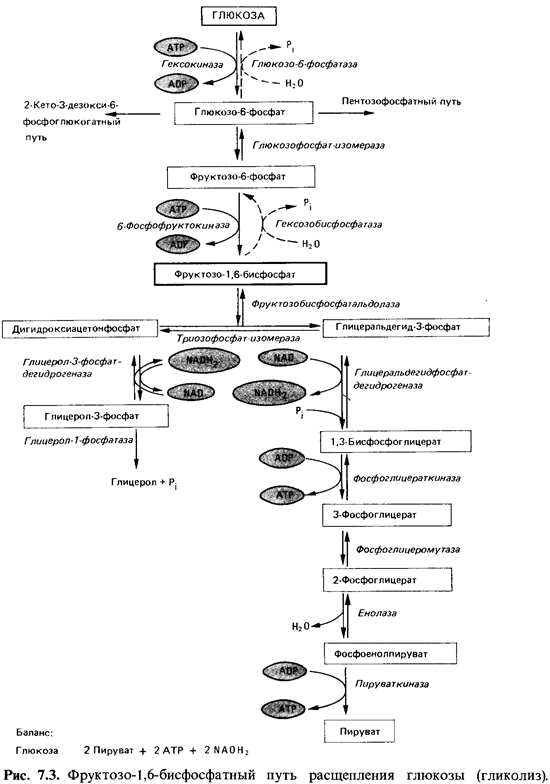
Другой ряд реакций,
к осуществлению которых способно большинство организмов, образует цикл,
известный под названием окислительного пентозофосфатного пути,
гексозомонофосфатного пути или схемы Варбурга-Диккенда-Хореккера (рис. 7.4).
Обратная последовательность; реакций этого пути включает важные
этапы, ведущие к регенерации акцептора СО2 при автотрофной фиксации
углекислоты. Только у бактерий встречается, видимо, путь Энтнера-Дудорова, или,
как его еще называют, КДФГ-путь (по характерному промежуточному
продукту-2-кето-3-дезокси-6-фосфоглюконату, КДФГ; рис. 7.5). Другие сходные
механизмы распада гексоз имеют более специальное значение. |
На фруктозодисфосфатном
пути (рис. 7.3) глюкозо-6-фосфат при подготовке к расщеплению изомеризуется глюкозофосфат-изомеразой в
фруктозо-6-фосфат; затем происходит фосфорилирование в положении 1 под
действием фосфофруктокиназы за счет АТР. Образовавшийся фруктозе-
1,6-дисфосфат расщепляется фруктозодисфосфатальдолазой до дигидроксиацетонфосфата
и глицеральдегид-3-фосфата. Оба триозофосфата находятся в равновесии между
собой; установление этого равновесия катализируется триозофосфат-изомеразой. Дигидроксиацетонфосфат
может восстанавливаться глицеролфосфат-дегидрогеназой до глицеролфосфата,
который гидролизуется глицерол-1-фосфатазой с образованием глицерола и
ортофосфата. Обычно же сначала происходит превращение образовавшегося под
действием альдолазы дигидроксиацетонфосфата в глицеральдегид-3-фосфат, который
затем окисляется. |
В
пентозофосфатном пути (рис. 7.4) глюкозо-6-фосфат дегидрируется глюкозо-6-фосфат-дегидрогеназой;
при этом водород переносится на NADP и образуется 6-фосфоглюконолактон, который
спонтанно или при участии фермента (глюконолактоназы) гидролизуется до
6-фосфоглюконата. Этот последний дегидрируется дегидрогеназой до З-кето-6-фосфоглюконата,
из которого затем путем декарбоксилирования образуется рибулозо-5-фосфат. Этим
завершается собственно процесс окисления. |
|
 |  |


