Дрожжи — одноклеточные существа, почти не способные к обучению в прямом смысле этого слова. Однако, как выяснилось, обычная дарвиновская эволюция, основанная на мутациях и отборе, способна отчасти компенсировать этот недостаток, помогая дрожжам вырабатывать полезные навыки, похожие на условные рефлексы животных. Фото с сайта www.bio.miami.edu Израильские ученые обнаружили у микроорганизмов
(кишечной палочки Escherichia coli и дрожжей Saccharomyces cerevisae)
способность к опережающему реагированию, напоминающую классические павловские
условные рефлексы. Если в естественной среде обитания микробов один стимул
часто предшествует другому, то микробы могут научиться реагировать на первый
стимул как на сигнал, предупреждающий о скором появлении второго стимула.
В отличие от собак Павлова, микробы приобретают свои «рефлексы» не путем
обучения, а за счет мутаций и отбора в длинной череде поколений.
Многие животные способны уловить в череде сигналов,
поступающих из внешней среды, определенные закономерности и использовать их для
«предвосхищения» грядущих событий — опережающего реагирования. В этом
суть классических условных рефлексов, изученных И. П. Павловым. Если
во время кормления собаки (или за несколько секунд до его начала) включать
звуковой сигнал, то вскоре одного этого сигнала будет достаточно, чтобы
у собаки началось слюноотделение.
У организмов, лишенных нервной системы, в том
числе у одноклеточных, способности к прижизненному обучению резко
ограничены. Но это не значит, что они принципиально не способны к опережающему
реагированию. Теоретически, они могут научиться предвосхищать события не хуже
павловских собак, но только не за счет прижизненного обучения, а за счет
эволюции. Иными словами, вместо «обычной» памяти, которая записывается в
структуре межнейронных связей, можно использовать память генетическую,
записанную в ДНК. Благодаря гигантской численности популяций микробов,
высокой скорости мутирования и очень быстрой смене поколений такое
«эволюционное обучение» у микробов теоретически может быть вполне
сопоставимо по своей скорости с «обычным» обучением у высших
животных.
Как и в случае с собаками Павлова, необходимым
условием для развития опережающего реагирования у микробов является
известная степень предсказуемости среды. Если стимул А всегда или очень
часто предшествует стимулу Б, и при этом природа стимула Б
такова, что к нему лучше начинать готовиться заранее, то организм может
научиться воспринимать стимул А как «предупреждение» о скором появлении стимула Б.
В такой ситуации естественный отбор может поддержать мутации, которые
позволят организму начать подготовку к стимулу Б в ответ на
«предупреждающий» стимул А.
Теоретически, в зависимости от того, каким образом два
стимула сочетаются друг с другом во времени, могут оказаться выгодными
разные стратегии реагирования (см. рисунок). Из четырех вариантов, показанных
на рисунке, первый — самый тривиальный — давно известен и изучен на
бесчисленном множестве примеров. Второй изучен хуже, но его существование
убедительно показано в экспериментах на дрожжах (Acar et al., 2008. Stochastic
switching as a survival strategy in fluctuating environments).
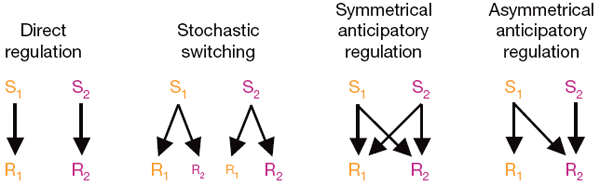
Четыре варианта реагирования на два стимула (S1 и S2):
1) прямая регуляция: каждый стимул вызывает только «свою» специфическую реакцию (это выгодно в том случае, если между стимулами нет никакой связи);
2) случайное переключение: разные особи в популяции реагируют на один и тот же стимул по-разному (это может повысить шансы выживания популяции в малопредсказуемой, быстро меняющейся среде);
3) симметричная опережающая регуляция: каждый из двух стимулов провоцирует сразу обе ответные реакции (выгодно, когда два стимула часто сближены во времени, но конкретная последовательность их появления заранее не известна);
4) асимметричная опережающая регуляция: первый стимул вызывает сразу обе реакции, а второй — только «свою» (выгодно, если первый стимул всегда «предвещает» приход второго, но не наоборот). Рис. из обсуждаемой статьи в Nature На третьем варианте (симметричное опережающее
реагирование) нужно остановиться подробнее. Он был открыт только в прошлом
году у кишечной палочки (Tagkopoulos et al., 2008. Predictive
behavior within microbial genetic networks). При этом впервые была
показана способность микробов к опережающему реагированию. Кроме того, впервые
удалось продемонстрировать, что эволюция генно-регуляторных сетей
у микроорганизмов действительно может имитировать процесс обучения у высших
животных.
Кишечная палочка живет попеременно то во внешней
среде (например, в воде или почве), то в пищеварительном тракте
млекопитающих. Когда бактерия попадает в желудок млекопитающего, она
сталкивается с двумя новыми стимулами: повышением температуры и нехваткой кислорода.
Оказалось, что у кишечной палочки выработалось «симметричное опережающее
реагирование» на эти стимулы. В ответ на повышение температуры
у бактерии активизируются не только гены, необходимые для защиты от
«теплового шока», но также и гены, необходимые для выживания в бескислородных
условиях. Это происходит даже в том случае, если кислорода в среде
пока еще много. Если подвергнуть бактерию кислородному голоданию, не повышая
при этом температуру среды, то бактерия тоже выдает сразу обе реакции: «прямую»
на отсутствие кислорода и «опережающую» — на тепловой шок, который пока
еще не наступил.
Бактерия как будто заранее знает, что если она попала
в тепленькое местечко, то скоро ей станет нечем дышать, а если ей стало нечем
дышать, то скоро, скорее всего, станет очень тепло. Эти выработавшиеся
в ходе эволюции «рефлексы» могут быть утрачены за несколько сотен
поколений, если бактерий выращивать в среде, где присутствует только один
из двух стимулов (например, в теплой среде с избытком кислорода). Точно
так же и собака Павлова постепенно перестает реагировать на звуковой сигнал,
если он больше не подкрепляется пищевым вознаграждением. Напомним, что
у бактерий, в отличие от собаки, «обучение» происходит за счет
мутаций и отбора и требует смены множества поколений.
Утрата бактериями опережающего реагирования при
разрыве связи между стимулами показывает, что сопряженность двух реакций (на
тепло и на кислородное голодание) не является каким-то базовым, фундаментальным
свойством генно-регуляторной сети бактерии. Две реакции на два разных
стимула — это действительно два разных процесса, которые могут в ходе
«эволюционного обучения» стать согласованными, а могут и утратить
согласованность, если в этом больше нет необходимости.
Таким образом, из четырех показанных на рисунке
вариантов неоткрытым у микробов оставался только последний: асимметричное
опережающее реагирование. Именно этот пробел и восполнили израильские ученые
в статье, опубликованной на сайте журнала Nature. Объекты для
исследования были выбраны классические: те же дрожжи и та же кишечная палочка.
Оба эти организма в течение своей жизни сталкиваются с резкими изменениями
условий. Самое главное, что временная последовательность этих изменений отчасти
предсказуема. Именно это и делает возможным развитие асимметричного
опережающего реагирования.
Авторы сформулировали три критерия, которым должно
соответствовать поведение микроба, чтобы его можно было классифицировать как
асимметричное опережающее реагирование:
1) Поведение должно быть
адаптивным, то есть выгодным. Иными словами, предварительное воздействие
стимула А должно повышать жизнеспособность микробов позже, когда начнет
действовать стимул Б. Это означает, что микробы действительно «уловили»
закономерность в последовательном появлении двух стимулов и научились
использовать стимул А как сигнал, по которому нужно начинать готовиться к
стимулу Б. Обратного эффекта быть не должно, то есть предварительное
воздействие стимула Б не должно повышать жизнеспособность
в условиях А. В противном случае это будет симметричное
опережающее реагирование, уже открытое ранее.
2) Подготовка к стимулу Б
должна быть дорогостоящей. Иными словами, заблаговременное включение генов,
необходимых для выживания в условиях Б, не должно приносить никакой пользы
до тех пор, пока условия Б реально не наступят. Это докажет, что
отбор поддержал опережающее реагирование не потому, что оно выгодно уже
в условиях А, а потому, что «убытки», понесенные микробом
в условиях А, будут с лихвой компенсированы той выгодой, которую
он получит позже, когда наступят условия Б.
3) Опережающая реакция должна
быть специфичной. Иными словами, микроб должен начинать «готовиться» к
условиям Б только в ответ на стимул А, который в естественной
среде обитания микроба обычно предшествует наступлению условий Б. Другие
стимулы не должны вызывать у микроба такой «подготовительной» реакции.
В противном случае эту реакцию можно будет интерпретировать иначе,
например как некий «обобщенный» ответ на стрессовые воздействия.
В экспериментах с кишечной палочкой авторы использовали
в качестве стимула А присутствие в среде сахара лактозы, а
в качестве стимула Б — присутствие другого сахара, мальтозы.
Дело в том, что в ходе своего путешествия по пищеварительному тракту
кишечная палочка сначала попадает в среду, где много лактозы, а затем
оказывается в среде, где много мальтозы. Поэтому теоретически бактерия
могла в ходе эволюции научиться воспринимать лактозу как сигнал,
предвещающий скорое появление мальтозы.
Эксперименты полностью подтвердили это предположение.
Оказалось, что в присутствии лактозы у бактерий активируются не
только гены, необходимые для усвоения лактозы, но и гены, требующиеся для
переваривания мальтозы. В присутствии мальтозы активируются только
«мальтозные» гены. В среде, где нет ни одного из двух сахаров, обе группы
генов остаются неактивными.
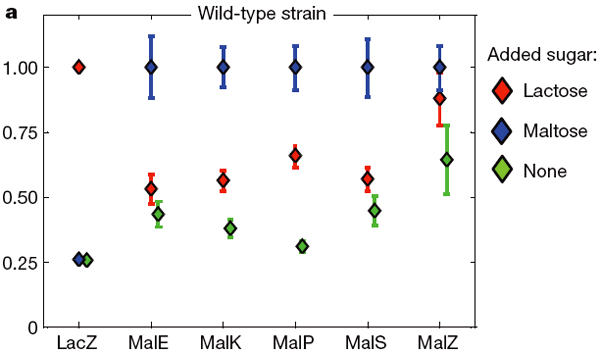
Активность «лактозных» (LacZ) и «мальтозных» (Mal*) генов у дрожжей при добавлении в среду лактозы (красные ромбики), мальтозы (синие) и в контрольной ситуации без обоих сахаров (зеленые). За единицу принята активность лактозных генов при добавлении в среду лактозы и мальтозных — при добавлении мальтозы. Видно, что лактоза активизирует работу мальтозных генов (красные ромбики выше зеленых в пяти правых столбцах), однако мальтоза не влияет на активность лактозного гена (синий ромбик на одном уровне с зеленым в левом столбце). Рис. из обсуждаемой статьи в Nature Чтобы доказать, что включение мальтозных генов
в ответ на лактозу действительно является асимметричным опережающим
реагированием, авторы последовательно применили три вышеперечисленных критерия.
Адаптивность оценивали по скорости размножения
бактерий в среде с мальтозой в течение первых двух часов после
помещения микробов в эту среду. Оказалось, что предварительное
«знакомство» с лактозой заметно повышает скорость размножения микробов в
мальтозной среде. Значит, бактериям действительно выгодно заранее включать
гены, необходимые для переваривания мальтозы (если они точно знают, что
мальтоза скоро будет подана к столу). Обратный эффект выявлен не был:
предварительное содержание в мальтозной среде нисколько не повышало
выживаемость микробов при последующем переводе в лактозную среду.
«Затратность» опережающего реагирования была доказана
при помощи эволюционного эксперимента. Авторы рассуждали примерно так.
Допустим, что «мальтозные» гены действительно совсем не нужны для усвоения
лактозы. Тогда их опережающее включение в среде с лактозой будет выгодно
только в том случае, если после лактозы в среде обязательно
появляется мальтоза. Если же мальтозы не будет, работа этих генов окажется
только лишней тратой ресурсов. Следовательно, если постоянно кормить микробов
одной лишь лактозой, отбор будет благоприятствовать таким бактериям-мутантам,
у которых механизм опережающего реагирования вышел из строя.
Проведенный эксперимент полностью подтвердил это
предположение. Бактерии, которых в течение 500 поколений выращивали в
лактозной среде без добавления мальтозы, полностью «разучились» включать свои
мальтозные гены в ответ на присутствие лактозы. Интересно, что при этом
они вовсе не утратили способность переваривать мальтозу. Мальтозные гены
сохранились в целости, и по-прежнему включались в ответ на появление
в среде самой мальтозы. Исчезло только опережающее реагирование.
По-видимому, это означает, что система усвоения мальтозы и прямая регуляция
этой системы более основательно «вмонтированы» в геном кишечной палочки, так что
для выхода их из строя требуется большее количество мутаций, чем для поломки
системы опережающего реагирования.
Специфичность реакции тоже была подтверждена в ряде
экспериментов. Например, было показано, что другие сахара (сахароза и
галактоза), в отличие от лактозы, не включают у кишечной палочки
мальтозные гены и не повышают ее выживаемость при переводе в среду, где
единственной пищей является мальтоза.
Любопытно, что бактерии, полученные в ходе
эволюционного эксперимента, у которых механизм опережающего реагирования
вышел из строя, при резком переводе из лактозной среды в мальтозную вообще не
успевают приспособиться к перемене и погибают.
Таким образом, кишечная палочка в ходе эволюции
действительно научилась воспринимать лактозу не только как пищу, но и как
сигнал, предвещающий скорое появление мальтозы.
В жизни дрожжей, как и у кишечной палочки,
некоторые изменения условий среды следуют друг за другом в строгом порядке, что
создает предпосылки для развития опережающего реагирования. Когда дрожжевые клетки
попадают в виноградный сок (в процессе изготовления вина) или просто на
гниющую виноградину, то они сначала начинают быстро размножаться, осуществляя брожение — быстрое и весьма «неэкономное»
переваривание сахаров без участия кислорода. В ходе брожения
вырабатывается много тепла. Среда может так сильно разогреться, что это
начинает угрожать жизни дрожжей. Справившись с тепловым шоком, дрожжи вскоре сталкиваются
с новой опасностью, исходящей от накапливающегося в среде этилового
спирта — побочного продукта брожения. Как и тепловой шок, высокая
концентрация спирта требует от дрожжей определенной защитной реакции,
то есть включения определенных генов. Вскоре наступает и третье
«потрясение»: в среде заканчиваются пригодные для сбраживания сахара, и
дрожжам приходится переключаться на более экономный, но и более опасный для них
способ питания — на окисление органики при помощи кислорода. Иными словами,
дрожжи начинают дышать. Для нас это единственно возможный вариант метаболизма,
а для дрожжей — дополнительный и далеко не оптимальный, к которому
дрожжи прибегают лишь в крайнем случае. В процессе дыхания в
митохондриях дрожжей образуется много свободных радикалов — это ведет коксидативному (окислительному) стрессу, с которым
тоже нужно бороться путем включения специальных генов.
Таким образом, три стрессовых воздействия следуют друг
за другом в строгом порядке: сначала тепловой шок, затем избыток спирта и,
наконец, оксидативный стресс. Каждый последующий стресс может быть «предсказан»
на основе предыдущих. Научились ли дрожжи использовать эту информацию? Серия
экспериментов, проведенных авторами, позволила дать положительный ответ на этот
вопрос. В каждом эксперименте дрожжи последовательно подвергались
воздействию двух разных стрессоров. Авторы исходили из того, что если
у дрожжей, как и у кишечной палочки, развилось асимметричное
опережающее реагирование, то «ранние» стрессоры (тепловой шок и спирт) должны
помогать дрожжам заблаговременно подготовиться к «позднему» оксидативному
стрессу. Обратного эффекта наблюдаться не должно или он должен быть гораздо
слабее.
Эти ожидания полностью подтвердились. Дрожжи,
пережившие оксидативный стресс, не стали от этого более устойчивыми к
нагреванию и отравлению спиртом. Напротив, дрожжи, пережившие тепловой шок или
побывавшие в среде с избытком спирта, во много раз лучше выдерживали
оксидативный стресс по сравнению с «наивными», неподготовленными дрожжами.
Таким образом были показаны адаптивность и
асимметричность опережающего реагирования у дрожжей. Авторы также
показали, что другие стрессоры — например, осмотический шок, высокая
концентрация меди или уксусной кислоты — не повышают устойчивость к
оксидативному стрессу. Тем самым была подтверждена специфичность реакции.
Затем авторы попытались выяснить, какие гены участвуют
в подготовке дрожжей к оксидативному стрессу в ответ на тепловой шок. Для
этого они измеряли активность (экспрессию) всех генов у дрожжей, которых
сначала подвергали нагреванию, а затем оксидативному стрессу. Для сравнения
использовались дрожжи, которых сначала подвергали осмотическому стрессу, а
затем оксидативному. Осмотический стресс, в отличие от теплового шока, не
вызывает у дрожжей «превентивной» защитной реакции. В результате
удалось выявить около 300 генов, которые нужны для защиты от оксидативного
стресса и которые включаются «заранее» при тепловом шоке, но не включаются при
осмотическом стрессе. Теперь нужно было доказать, что эти гены не нужны для
защиты от теплового шока и что их включение при тепловом шоке является
превентивной мерой — подготовкой к оксидативному стрессу, который
ожидается в будущем, но пока еще не начался. Для этого авторы использовали
29 штаммов дрожжей, у каждого из которых был удален один ген из числа
тех 300, что были выявлены на предыдущем этапе исследования. Все эти
штаммы проверили на устойчивость к тепловому шоку и оксидативному стрессу.
Оказалось, что 65% штаммов отличаются пониженной
устойчивостью к оксидативному стрессу. Однако тепловой шок все они переносят
ничуть не хуже, чем контрольные «дикие» дрожжи. Следовательно, эти гены
действительно нужны для защиты от оксидативного стресса, но они не нужны для
борьбы с тепловым шоком. Включение этих генов при тепловом шоке не может быть
ничем иным, кроме как опережающим реагированием — подготовкой к
приближающемуся оксидативному стрессу. При этом тепловой шок воспринимается
дрожжами как сигнал о том, что скоро им придется дышать и что пора готовиться к
борьбе со свободными радикалами.
Таким образом, наличие у микроорганизмов способности к
асимметричному опережающему реагированию можно считать доказанным. При
определенной степени предсказуемости изменений окружающей среды эволюция
генно-регуляторных сетей может приводить к результатам, очень похожим на
выработку условных рефлексов у животных, когда одно событие
интерпретируется как предвестник другого. Пока остается открытым вопрос о том, насколько
широко распространено такое «эволюционное обучение» в природе.
Источник: Amir Mitchell, Gal H. Romano, Bella Groisman, Avihu Yona, Erez Dekel, Martin Kupiec, Orna Dahan, Yitzhak Pilpel. Adaptive prediction of environmental changes by microorganisms // Nature. Advance online publication 17 June 2009. Doi:10.1038/nature08112. |




