
Рис. 1. Многоклеточные трихомы (нити) цианобактерий Oscillatoria ползают друг по другу, иногда образуя интересные «супраклеточные» структуры, например плотно сплетенные «коврики» и разнообразные тяжи. Каждый трихом представляет собой «стопку» из множества клеток, формой напоминающих монету. Фото с сайта www.dr-ralf-wagner.de Нитчатые цианобактерии способны к скользящему
движению. Движение каждой нити подчиняется простым правилам, следование которым
может приводить к самосборке сложных структур из множества нитей, в том
числе туго скрученных, прочных «веревок». Анализ распространения цианобактерий,
вьющих такие веревки, и свойств субстратов, на которых они обитают,
показал, что данная способность, по-видимому, помогает микробам обживать
сыпучие, неустойчивые грунты.
Нитчатые цианобактерии — одни из самых
«высокоорганизованных», то есть сложно устроенных, прокариот. Нить, или
трихом, формируется в результате не доведенных до конца клеточных делений
и состоит из множества расположенных в один ряд клеток. У некоторых видов
наблюдается разделение функций между клетками трихома: одни клетки
специализируются на фотосинтезе, другие — на фиксации азота (эти процессы
трудно совместить в одной клетке, хотя недавно выяснилось, что некоторым
цианобактериям это удается, см.: Цианобактерии совмещают в одной клетке фотосинтез и фиксацию
атмосферного азота, «Элементы», 01.02.2006).
Нитчатые цианобактерии со специализацией клеток —
одна из наиболее далеко зашедших «попыток» эволюции создать многоклеточный
организм на прокариотной основе. Впрочем, усложнение таких нитчатых
«многоклеточных организмов» так и не продвинулось дальше самых первых шагов.
Однако некоторые нитчатые цианобактерии перешли к следующему уровню
интеграции — формированию упорядоченных структур из множества трихомов.
Это достигается благодаря способности трихомов к медленному скользящему
движению.
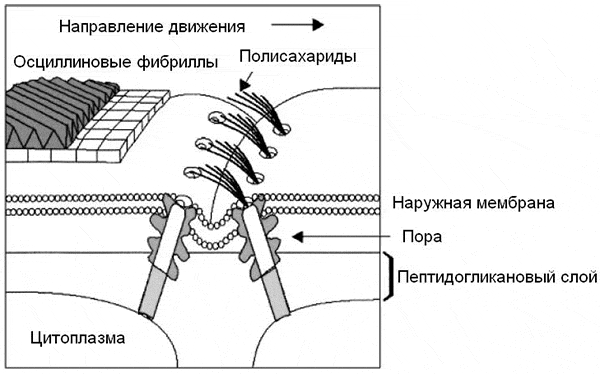
Рис. 2. Один из механизмов скользящего движения нитчатых цианобактерий. На рисунке показан участок трихома с границей между двумя клетками. Молекулы полисахаридов выталкиваются из специальных пор в направлении, противоположном направлению движения. Из-за косо расположенных осциллиновых фибрилл трихом может в ходе движения вращаться вокруг своей продольной оси. Рис. из статьи Mark J. McBride, 2001. Bacterial gliding motility: Multiple mechanisms for cell movement over surfaces Способ движения, показанный на рис. 2
(при помощи направленного выталкивания молекул полисахаридов через поры в
клеточной стенке), может показаться крайне расточительным, но полисахариды не
пропадают зря: из них постепенно образуется общий слизистый чехол,
защищающий сообщество трихомов от высыхания и прочих невзгод.
Ползая друг по другу, трихомы могут сплетаться в
плотные «коврики», покрывающие поверхность субстрата. Наблюдение за поведением
цианобактерий — увлекательное занятие. Например, если такой коврик
разорвать, из краев разрыва вскоре высовываются отдельные трихомы, которые,
постепенно выползая всё дальше, начинают совершать круговые «ищущие» движения.
Наткнувшись на ползущего навстречу (из противоположного края разрыва) товарища,
трихом прилипает к нему, и они начинают ползти друг по другу в противоположные
стороны. Поскольку каждый трихом, по-видимому, прочно «заякорен» в своей
половине коврика, это создает усилие, стремящееся соединить разъединенные
половинки. Многочисленные пары ползущих друг по другу трихомов действуют
подобно сокращающимся мышечным волокнам. В конце концов половинки коврика
приходят в движение, края разрыва сближаются, и трихомы быстро «заплетают»
разрыв (см.: Е. Л. Сумина, 2007. Экспериментальное изучение сообщества нитчатых цианобактерий,
PDF, 1,03 Мб; я имел счастье понаблюдать за поведением нитчатых
цианобактерий Oscillatoriaв связи с тем, что был оппонентом на защите
этой диссертации и попросил диссертанта принести мне культуру).
Такое сложное и «осмысленное» коллективное поведение
наводит некоторых авторов на раздумья, а нельзя ли организованное сообщество
нитчатых цианобактерий считать настоящим целостным организмом? По-моему, это
слишком громко сказано, и таких терминов, как «коллективное поведение» и
«организованное сообщество» здесь вполне достаточно (см.: «Чувство кворума»:
принятие коллективных решений в макро- и микромире, «Элементы»,
02.04.2009). Однако какой-то специальный термин для обозначения сложных
структур, образованных множеством многоклеточных трихомов, все-таки нужен.
Некоторые авторы называют такие структуры «супраклеточными».
Одной из типичных супраклеточных структур являются
прочные, туго скрученные «веревки», покрытые общим полисахаридным чехлом
(рис. 3). Способность плести веревки характерна для родов Microcoleus, Schizotrix иHydrocoleum.
Эту способность использовали при классификации цианобактерий в качестве
одного из таксономических признаков. То есть предполагалось, что она
унаследована бактериями от общего предка, который первым ее приобрел и передал
потомкам.
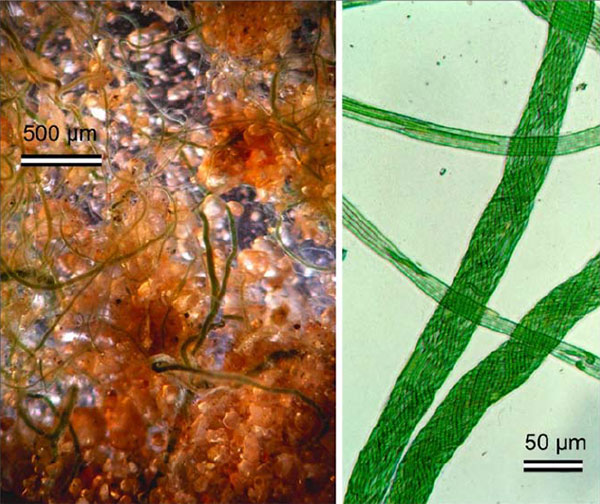
Рис. 3. Слева: поселение цианобактерий Microcoleus на рыхлом песчаном грунте (в пустыне в штате Колорадо); хорошо видны отдельные «веревки», оплетающие песок. Справа: «веревки» при большом увеличении; видно, что каждая веревка сплетена из множества трихомов. Трихомы микроколеуса гораздо тоньше, чем у изображенной на рис. 1 осциллатории, которая настоящих веревок не плетет. Фото из обсуждаемой статьи в PLoS ONE Генетические основы «плетения веревок» неизвестны,
однако есть все основания полагать, что это действительно наследственный,
генетически детерминированный признак. Это видно, например, из того, что
в природе виды, вьющие веревки, встречаются по большей части именно
в виде веревок, а при культивировании в лаборатории они иногда вдруг
теряют эту способность безвозвратно — видимо, в результате каких-то
мутаций.
Каждый раз, когда биологи сталкиваются с необычным
признаком, перед ними встает классический «дарвиновский» вопрос: зачем этот
признак нужен? Конечно, далеко не всякий признак имеет адаптивное значение:
многие особенности строения и поведения живых организмов закрепляются
в ходе эволюции не под прямым действием отбора, как приносящие пользу, а
случайно, из-за стечения исторических обстоятельств, или как побочный продукт
других изменений. Тогда правильнее спрашивать не зачем появился
признак, а просто почему. Что касается веревкоплетения у цианобактерий, то
до сих пор никто не знал (или, по крайней мере, всерьез не
задумывался), какой из двух вопросов в данном случае уместнее.
Исследователи из Университета штата Аризона решили разобраться в этой
«запутанной» проблеме. Они рассудили, что плетение веревок должно давать
бактериям какие-то преимущества хотя бы потому, что вред такого
поведения вполне очевиден. Для отдельного трихома жизнь в составе веревки
сопряжена с целым рядом серьезных неудобств по сравнению с независимым
существованием. Скрученные в толстые жгуты трихомы загораживают друг другу
свет, что затрудняет фотосинтез — это неудобство становится ощутимым
начиная с толщины веревок около 10 мкм (а их реальная толщина достигает
50–100 мкм). Изолированному трихому намного легче получать из окружающей
среды необходимые вещества и избавляться от отходов жизнедеятельности (одним из
главных отходов жизнедеятельности цианобактерий является, между прочим, молекулярный
кислород). Эти неудобства должны компенсироваться какой-то существенной
выгодой, иначе отбор давно отбраковал бы мутации, изменившие поведение бактерий
таким образом, что они стали вить из себя веревки.
Начали исследователи с того, что собрали данные по
нуклеотидным последовательностям двух генов (16S рРНК и kaiC) у
30 природных и лабораторных штаммов цианобактерий, вьющих веревки,
а также у множества других цианобактерий из всех основных подразделений этой
обширной группы. На сегодняшний день прочтены геномы 36 цианобактерий
(см.:Complete
Microbial Genomes), но «веревочных» видов среди них нет. Поэтому выбор
генов для анализа определялся в первую очередь наличием доступных данных
по «веревочным» разновидностям.
На основе этих нуклеотидных последовательностей были
построены эволюционные деревья. Это делалось, чтобы выяснить, имеют ли формы,
умеющие свиваться в веревки, единое происхождение. Но анализ показал, что
плетение веревок появилось в эволюции цианобактерий как минимум четыре раза
независимо. Это видно из того, что «веревочные» штаммы образуют на дереве
четыре компактные ветви, раскиданные по самым дальним концам дерева и никак
друг с другом не связанные. Любопытно, что все эти штаммы, выделенные цветом на
рис. 4, формально (то есть согласно принятой классификации) относятся
к одному и тому же роду Microcoleus. Этот пример показывает, чего стоят
старые классификации микробов, основанные на внешних признаках, в свете
новых молекулярных данных. Но микробиологи уже привыкли к этому и не
удивляются, и даже не торопятся переименовывать свои объекты (по-хорошему, надо
было бы разделить род Microcoleus на четыре рода, но это создаст ужасную
путаницу, и поэтому никто не спешит вносить коррективы в привычную
номенклатуру). На том же дереве можно найти еще один такой несчастный «род», на
деле оказавшийся произвольным собранием неродственных, хотя и похожих внешне
микробов (Synechococcus).
В принципе, можно было бы объяснить такое
распределение «веревочных» штаммов тем, что способность плести веревки
появилась очень давно у их далекого общего предка и была впоследствии утрачена
другими потомками этого предка. Но тогда пришлось бы признать, что предками
чуть ли не всех современных цианобактерий, в том числе одноклеточных, были
нитчатые формы, плетущие веревки. Такое допущение кажется, мягко говоря,
сомнительным. Поэтому авторы вполне законно останавливаются на версии о
многократном независимом появлении способности плести веревки у разных
нитчатых цианобактерий.
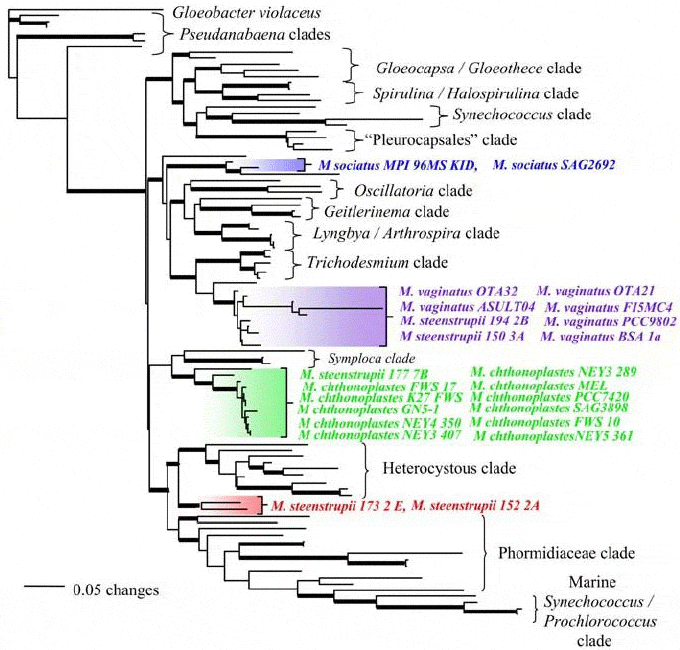
Рис. 4. Эволюционное древо цианобактерий, основанное на гене 16S рРНК. Формы, умеющие вить веревки, образуют четыре изолированные ветви (выделены цветом). Рис. из обсуждаемой статьи в PLoS ONE Многократное появление признака — веский довод
в пользу его полезности. Эволюция может много раз «натыкаться» на одно и
то же конструктивное решение и закреплять его, если оно удачно, но всевозможные
«эволюционные случайности» обычно происходят (точнее, закрепляются) только
единожды. Примером такой случайности считается, в частности, пятипалая конечность наземных
позвоночных.
Итак, теперь можно было с уверенностью ставить
вопрос «зачем». Авторы проанализировали все имеющиеся литературные данные по
«веревочным» цианобактериям, а также результаты собственных наблюдений,
пытаясь найти что-то общее в их образе жизни. Эти формы живут в очень
разных местах: на песчаных морских отмелях в приливно-отливной зоне, на рыхлых
карбонатных грунтах на мелководьях и даже в пустынях на голом песке. Тем
не менее авторам удалось выделить три экологические особенности, общие для всех
«веревочных» штаммов:
1) Они всегда живут на самых
рыхлых, подвижных, неустойчивых грунтах, наиболее уязвимых для эрозии.
2) Они обычно выступают в роли
пионеров — первопоселенцев на этих грунтах, обживая их раньше всех прочих
живых существ, в том числе других прокариот (см.: Цианобактерии —
первопоселенцы высокогорных пустынь, «Элементы», 10.09.2008).
3) Они очень эффективно
скрепляют частицы грунта, защищая его от эрозии. Многие прокариоты делают то же
самое, склеивая песчинки выделяемыми органическими веществами или цементируя их
при помощи биоминерализации, но мало кто из них может сравниться по
эффективности укрепления сыпучего грунта с «веревочными» цианобактериями.
Эти факты явно указывают на то, что толстые
«супраклеточные» веревки, покрытые липким полисахаридным чехлом, по-видимому,
представляют собой оптимальный способ организации трихомов, позволяющий
наиболее эффективно укреплять самые неустойчивые разновидности субстратов —
и тем самым делать их пригодными для жизни.
В качестве дополнительного аргумента, подкрепляющего
это предположение, авторы приводят график зависимости устойчивости осадочных
пород к эрозии (эта устойчивость измеряется как скорость ветра или течения,
достаточная для отделения частиц от субстрата) от диаметра частиц. Этот график,
основанный на обширных литературных данных по седиментологии (см. Sedimentology),
имеет V-образную форму (рис. 5). При размере частиц менее 50–100 мкм
главную роль в скреплении грунта играют силы прилипания, или когезии (см.: Cohesion).
При этом чем мельче частицы, тем прочнее грунт. Наибольшая уязвимость
характерна для грунтов с диаметром частиц 50–100 мкм. При диаметре свыше
100 мкм главную роль начинает играть гравитация, то есть вес частиц.
При этом зависимость меняется на противоположную: чем крупнее частицы, тем
стабильнее грунт.
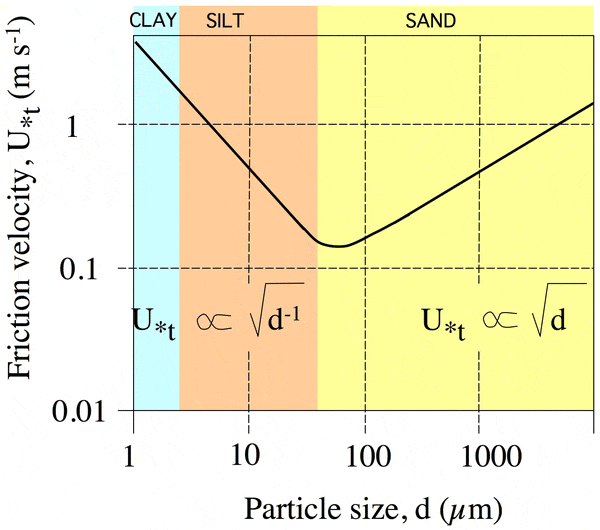
Рис. 5. Зависимость устойчивости пород к эрозии (вертикальная ось) от диаметра частиц (горизонтальная ось). Область перегиба на этом графике довольно точно соответствует толщине цианобактериальных «веревок» (50–100 мкм). Рис. из обсуждаемой статьи в PLoS ONE Авторы подчеркивают, что диаметр цианобактериальных
веревок довольно точно совпадает с «критическим» размером частиц грунта, при
котором субстрат наименее стабилен. Вероятно, преимущество толстых веревок по
сравнению с отдельными трихомами (или тонкими супраклеточными тяжами) состоит в
том, что уже с самого начала своего роста такие веревки гарантированно
выводят склеиваемые ими в единый конгломерат частицы грунта за пределы
нисходящей (левой) части графика и переносят их на правую, восходящую часть.
После этого чем длиннее веревка, тем надежнее ее минеральное «основание». Если
бы трихомы росли поодиночке, то их рост, сопровождающийся склеиванием очень
мелких частиц, поначалу мог бы приводить не к укреплению, а, наоборот, к
ослаблению грунта. Можно предложить и другие объяснения, почему «плетение
веревок» является оптимальной стратегией для нитчатых цианобактерий на сыпучих
грунтах. Например, лабораторные наблюдения показывают, что отдельные трихомы,
облепленные со всех сторон мелкими частицами грунта, чаще всего просто
погибают (им очень легко «испачкаться» со всех сторон, ведь они крутятся
вокруг продольной оси, когда ползают). Может быть, сплетаясь в веревки,
цианобактерии предохраняют себя от такой участи.
То, что «веревочные» цианобактерии обычно являются
пионерами — то есть поселяются на необжитых участках, скрепляют
грунт, а затем уступают место другим микробам, — хорошо согласуется с
тем упомянутым выше обстоятельством, что жизнь в виде веревок снижает
эффективность обмена веществ (фотосинтеза, извлечения веществ из окружающей
среды и удаления отходов). Пока «веревочные» формы живут на голом песке
в одиночестве, без всякой конкуренции, всё это можно перетерпеть. Но как
только на субстрате появляются более эффективные конкуренты, «веревки» начинают
проигрывать соревнование и вскоре оказываются вытеснены с обжитого места,
которое они сами же и сделали пригодным для жизни.
Источник: Ferran Garcia-Pichel, Martin F. Wojciechowski. The Evolution of a Capacity to Build Supra-Cellular Ropes Enabled Filamentous Cyanobacteria to Colonize Highly Erodible Substrates // PLoS ONE. 2009. 4(11): e7801. doi:10.1371/journal.pone.0007801. |




