Биохимическое единство, мысль о котором еще несколько десятилетий назад казалась столь невероятной, в настоящее время-твердо установленный факт. Клетки всех живых существ, от самых примитивных форм до наиболее высокоразвитых животных и растений, состоят из од них и тех же структурных элементов и используют одни и те же механизмы для получения энергии и для роста. По сравнению с этим фунда ментальным единством существующие различия и отклонения кажутся незначительными. Можно считать, что все ныне живущие организмы проделали вместе длинный путь развития. Из простейших форм посте пенно развились формы более сложные и специализированные, а потом, наконец, и те, которые населяют нашу планету сегодня. Этот процесс эволюционного развития организмов - одна из центральных проблем биологии. Первичная
атмосфера Земли. Наша Земля кардинальным образом отличается от других
планет Солнечной системы. По сравнению с Юпитером и Солнцем она содержит лишь
незначительные количества инертных газов. По-видимому, она образовалась в
результате объединения множества метеоритов; вследствие нагрева и расплавления
внутреннего ядра Земли вода и газы были вытеснены к ее поверхности. Первичная
атмосфера, вероятно, содержала много водорода, метана, азота и С02,
но в ней не было кислорода. При фотолизе водяных паров, разумеется,
освобождался кислород, но он вновь переходил в связанное состояние. Химическая
эволюция могла происходить только в бескислородной атмосфере.
Химическая
эволюция. Гипотеза, согласно которой жизнь была зане сена на нашу
планету извне, вряд ли заслуживает в настоящее время серьезного обсуждения.
Самовоспроизводящиеся биологические единицы должны были возникнуть на самой
Земле в ранний период ее существования. Согласно представлению, выдвинутому
Холдейном и Опариным, в то время на Земле накопились большие количества органических
веществ, но еще не было организмов, способных их использовать и минерализовать.
Когда после первых попыток Миллера уда лось неоднократно подтвердить в
эксперименте, что из неорганических веществ (Н2, С02, NH3,
Н20) и метана при подходящих условиях могут синтезироваться простые
органические молекулы, сомнения в реальности химической эволюции полностью
отпали. Как полагают, в восстановительной первичной атмосфере (в которой не
было кислорода) под действием солнечной радиации и в результате электрических
разрядов образовывались органические вещества, которые затем попадали в воду и
в ней накапливались. Когда они накопились в большом количестве, видимо,
возникли условия, при которых мог совершиться переход от химической эволюции к
возникновению первых самовоспроизводящихся живых существ.
Биологическая
эволюция. Переход от неживой органической материи к живой клетке
потребовал длительного времени (от 3,1 до 4,5 млрд. лет). Появившиеся клеточные
организмы получили, очевидно, столь большое селективное преимущество, что все
предшествующие формы организации оказались вытесненными. Поскольку доклеточные
формы жизни (если они существовали) не сохранились даже в ископаемом виде,
переход от неживого к живому представляется нам чрезвычайно быстрым.
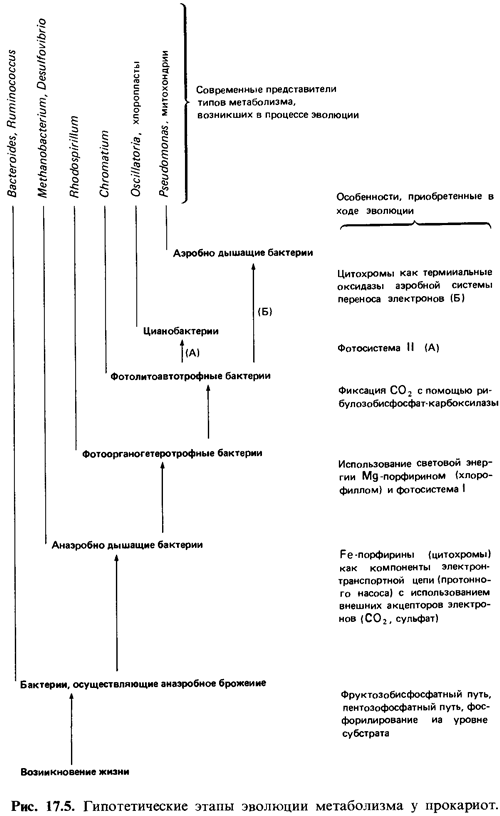
Эволюция
прокариот. Согласно распространенному, хотя и весьма гипотетическому
представлению, в восстановительной первичной атмосфере происходило развитие
прокариотических организмов (рис. 17.5). Первыми прокариотами, которые могли
появиться в водоемах, богатых органическими веществами, были организмы,
существовавшие за счет брожения и обладавшие основными функциями анаэробного
обмена (фруктозобисфосфатный и пентозофосфатный пути). Если предположить, что в
водоемах имелись тогда и сульфаты, то следующим достижением органической
эволюции мог быть эффективный транспорт электронов с созданием протонного
потенциала как источника энергии для регенерации АТР. На этом этапе эволюции,
вероятно, возникли производные тетрапиррола, содержащие железо или никель, а
также автотрофный способ ассимиляции углерода (путь ацетил-СоА). Как реликты
тех времен могут рассматриваться метанобразующие и ацетогенные бактерии, а
также бактерии, восстанавливающие сульфаты до сульфида, которые, за рядом
исключений, могут использовать Н2, СО2 и некоторые
продукты брожения.
После
«изобретения» фосфорилирования, сопряженного с переносом электронов, могла
возникнуть также фотосистема I - «протонный насос, приводимый в действие
светом», что позволило использовать свет в качестве источника энергии. Реакционными
центрами служили магнийпорфирины (хлорофиллы). Первые фототрофные организмы,
вероятно, ассимилировали углерод на свету подобно Rhodospirillaceae. С
приобретением способности фиксировать СО2 в
рибулозобисфосфатном цикле и использовать неорганические доноры электронов (Н2,
H2S, S) выработался тип метаболизма, характерный для пурпурных
серных бактерий (Chromatiaceae). К еще большей независимости от растворенных в
воде веществ привело затем появление фотосистемы II: стал возможен нециклический
перенос электронов с использованием воды в качестве их донора. Этот процесс был
неизбежно связан с выделением кислорода. Оксигенный фотосинтез привел к тому,
что земная атмосфера приобрела окислительный характер. Представителями первых
микроорганизмов, осуществлявших фотосинтез с выделением О2, являются
цианобактерии.
Переход от
первичной восстановительной атмосфры к атмосфере, содержащей
кислород, несомненно, был величайшим событием как в эволюции живых
существ, так и в преобразовании минералов. В результате превращения цитохромов
в терминальные оксидазы и использования молекулярного кислорода в качестве
акцептора электронов у бактерий стал возможным новый тип метаболизма - аэробное
дыхание.
Как полагают,
2,1 млрд. лет назад уже существовали все фототрофные дышащие прокариоты,
известные в настоящее время. Согласно геологическим данным, уже 2,7 млрд. лет
назад имелся в небольшом количестве кислород. На протяжении последних 1,2 млрд.
лет вся жизнь на Земле зависит от биологического фотосинтеза и от кислорода,
выделяемого растениями. Вызвав накопление кислорода в атмосфере, развитие жизни
тем самым - через окисление металлов и минералов повлияло и на неживую природу.
В период до
0,6 млрд. лет назад содержание кислорода в атмосфере увеличилось, вероятно,
всего лишь до 2%. И только после того, как растения завоевали
сушу и покрыли ее густым зеленым ковром, концентрация кислорода в воздухе резко
повысилась и достигла современного уровня (21%). Накопление О2 сопровождалось
образованием отложений углерода в форме каменного угля, нефти, природного газа
и углеродсодержащих осадочных пород.
Ископаемые
остатки, относящиеся к раннему докембрию, чрезвычайно редки. Из-за малой
величины примитивных организмов и отсутствия у них каких-либо твердых
компонентов остатки их могли сохраниться лишь при исключительных
обстоятельствах. В штате Миннесота (США) в отложениях, возраст которых
оценивают в 2,7 млрд. лет, были обнаружены структуры, интерпретируемые как остатки
бактерий (в том числе цианобактерий). Возраст южноафриканских отложений, в
которых тоже были найдены структуры, напоминающие бактерии, достигает 3,1 млрд.
лет. Это самые древние из всех известных следов жизни.
Бактерии - это
сохранившиеся до наших дней живые свидетели ран ней эволюции жизни. Многие в
прошлом широко распространенные и господствовавшие бактерии в настоящее время
ведут весьма скромное существование. В экологических нишах, обеспечивающих им
подходящие условия жизни, сохранились также и анаэробные бактерии.
Эволюция
эукариот. Эукариотические клетки, видимо, возникли лишь тогда, когда в
атмосфере появился кислород. Все эукариоты, за очень малым исключением, -аэробные
организмы. Прокариоты занимали много различных экологических ниш. Выработка разнообразных
типов метаболизма у прокариот была, по-видимому, обусловлена простой структурой
клетки, высокоразвитыми системами регуляции, быстрым ростом и наличием
нескольких механизмов переноса генов. На пути дальнейшей эволюции прокариот
стояли непреодолимые трудности, связанные прежде всего с малыми размерами
генома, его гаплоидным состоянием и малой величиной клеток. Новая окружающая
среда с аэробными условиями позволяла получать больше энергии, но для ее использования
нужны были более крупные клетки, широкие возможности структурной
дифференцировки и соответственно во много раз больший геном, который
обеспечивал бы хранение большого объема информации. Величина генома 5-109 Да
была, вероятно, верхним пределом молекулярной массы бактериальной хромосомы,
состоящей из одной-единственной двойной цепи. Для дальнейшей эволюции
требовалось создание новой модели.
Различия
между прокариотической и эукариотической клетками (эуцитом и
протоцитом) огромны. Еще раз перечислим важнейшие особенности клеток эукариот:
1. Носитель
наследственной информации (ДНК) отделен от «метаболических пространств» ядерной
оболочкой.
2. Вследствие
этого транскрипция (в ядре) отделена от трансляции (в цитоплазме).
3. Геном
разделен на части, имеется несколько (часто много) линейных хромосом вместо
одной кольцевой.
4. Репликация
ДНК происходит только в интерфазе; каждая хромосома имеет по нескольку
репликонов; дочерние хромосомы распределяются путем митоза.
5. Существуют
внутриклеточные механизмы с использованием актина и тубулина для перемещения
хромосом во время митоза и мейоза, а также структуры типа пузырьков (лизосомы,
пероксисомы и другие «микро тельца»).
6. В генах
имеются некодирующие вставки - интроны.
7. ДНК
образует комплекс с гистонами, по структуре напоминающий нитку жемчуга (цепь из
нуклеосом).
8. Жизненный
цикл включает мейоз, при котором из диплоидных клеток образуются гаплоидные.
Это позволяет осуществить половой процесс с перекомбинированием генов и смену
гаплофазы и диплофазы.
9. Экзоцитоз:
внеклеточные ферменты синтезируются не прямо на плазматической мембране (с
одновременным выведением их из клетки), а на внутренних мембранах, после чего в
цистернах доставляются на поверхность.
10. Эндоцитоз
(в форме фагоцитоза и пиноцитоза), позволяющий приобретать внутриклеточных
симбионтов.
11. Наличие
митохондрий и хлоропластов, которые служат для получения энергии (ресинтеза
АТР).
12. Жгутики
(или реснички) типа 9 + 2.
Итак, эуцит
отличается от протоцита многими функциями и структурами. Хотя и известны
отдельные эукариоты, у которых тот или иной признак отсутствует, нет таких
примитивных форм, по которым можно было бы определить, в какой
последовательности появлялись новые признаки. По-видимому, каждый этап эволюции
приносил с собой лишь очень небольшое селективное преимущество, по крайней мере
по сравнению с ближайшей предшествующей ступенью. Таким образом, промежуточные
формы не сохранялись и, вероятно, были такими нестойкими, что сейчас нет даже
ископаемых остатков, которые позволяли бы судить об их функциональных особенностях.
В настоящее время имеется лишь небольшое число организмов, которые можно
считать развившимися из промежуточных форм. Возможность когда-либо установить
последовательность появления перечисленных выше новых признаков следует оценить
пессимистически. Но все же можно предположить, что на ранних этапах эволюции
эукариотической клетки возникали различные модели ее организации, прежде чем
появились многоклеточные организмы.
Следует
отметить, что эукариоты специализировались в основном на фотосинтезе и
существовании в аэробных условиях, а целый ряд других важных экологических
функций остался за прокариотами. К ним относятся фиксация азота, нитрификация,
денитрификация, сульфатное и серное дыхание, окисление серы и металлов,
образование и использование метана. Круговорот азота и серы полностью или
преимуществен но находится «в ведении» прокариот. Таким образом, прокариоты могли
бы поддерживать круговороты веществ и сохранять биосферу, тогда как эукариоты
одни не справились бы с этой задачей.
Если прокариоты
в течение миллиардов лет развивались сами по себе, то эукариоты никогда не
оставались одни. Им приходилось все время противостоять прокариотам. Они
предоставляли последним новые экологические ниши, защиту и были их жертвами.
Многоклеточные организмы своими высокоразвитыми защитными и иными приспособлениями
отчасти обязаны агрессивности прокариот. С другой стороны, эукариоты научились
извлекать пользу из тесной ассоциации с прокариотами и поставили их себе на
службу в качестве эктосимбионтов (в кишечном тракте, на коже, у жвачных в
рубце) и эндосимбионтов (для фиксации азота, продукции биомассы путем
фотосинтеза, использова ния H2S, удаления Н2).
Эволюция
живых организмов предлагает для решения массу увлекательных проблем. Их
исследование только начинается.
| 



