Дезоксирибонуклеиновая
кислота (ДНК) - макромолекула. При кислотном гидролизе она расщепляется на свои
структурные элементы - дезоксирибозу, фосфорную кислоту и азотистые
основания в эквимолярных соотношениях. В состав ДНК входят четыре
различных основания: два пуриновых (аденин и гуанин) и два пиримидиновых
(цитозин и тимин). Если
произвести расщепление ДНК с помощью нуклеаз (ДНКазы I из
поджелудочной железы или диэстеразы змеиного яда), то освобождаются
3'- или 5'-нуклеотиды. Эти дезоксирибонуклеотиды соединены в нуклеиновых
кислотах в длинные цепи. В ДНК молекулы пентозы чередуются с молекулами
фосфорной кислоты; с каждым сахаром связано одно из четырех оснований. Такая
полинуклеотидная цепь не одинакова в обоих направлениях, т.е. обладает
полярностью: на одном ее конце находится фосфатная группа в положении 5', а на
другом - свободная гидроксильная группа в положении 3'.
Еще в 1950 г. Чаргафф установил
ряд закономерностей: аденин присутствует в ДНК в том же количестве, что и
тимин, а гуанин - в том же, что и цитозин (А = Т, G = С); сумма пуриновых
оснований равна сумме пиримидиновых оснований; отношение (G + С)/(А + Т) может
варьировать в широких пределах от вида к виду, но остается постоянным для
данного вида.
Пространственное
расположение отдельных компонентов (т.е. вторичная структура ДНК)
было выяснено с помощью рентгеиоструктуриого анализа. Если
вытянуть ДНК в нить и вращать ее в монохроматическом пучке рентгеновского
излучения, а отклоняющиеся лучи проецировать на пленку с эмульсией, то можно
получить дифракционную рентгенограмму. Рентгенограммы ДНК различного
происхождения (из спермы, тимуса, бактериальных клеток и бактериофагов)
оказались почти идентичными. Анализ таких рентгенограмм, произведенный
Уилкинсом, показал, что пуриновые и пиримидиновые кольца располагаются под
прямым углом к продольной оси полинуклеотидного тяжа; этот тяж дол жен быть
спирально закручен вокруг центральной оси с шагом 3,4 нм. Исходя из данных о
плотности, следовало предполагать, что спираль состоит более чем из одного тяжа. 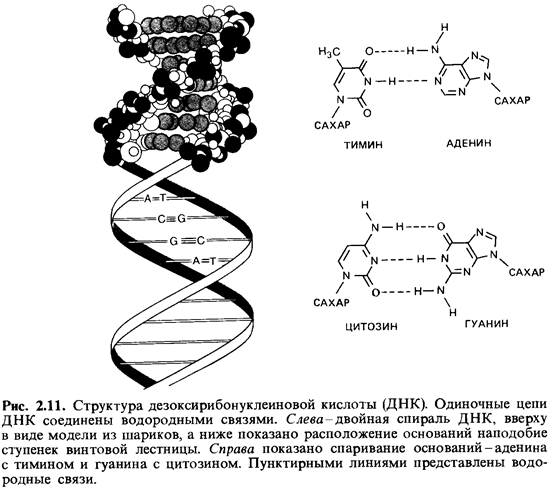
Полученные
результаты дали возможность Уотсону и Крику сформулировать (в 1953 г.) блестящую теорию о
структуре ДНК. Согласно модели Уотсона-Крика, полинуклеотидные цепи закручены в
двойную спираль вокруг воображаемой оси. Обе цепи скреплены водородными связями,
соединяющими основания, которые обращены внутрь спирали (рис. 2.11) При этом
если учесть расстояния и условия образования связи, то против каждого аденина
должен находиться тимин, а против каждого гуанина-цитозин. На один виток
спирали приходится около 10 пар оснований. Последовательность оснований в двух
цепях по необходимости комплементарна. Направление цепей противоположно (5'
-»3' и 3'-»5'). Длина хромосомы Escherichia coliсоставляет
примерно 1,4 мм.
Один микрометр одноцепочечной ДНК соответствует относительной молекулярной
массе около 2-106, или примерно 3000 пар оснований. Таким образом,
хромосома Е. coli имеет молекулярную массу 2,9-109. Водородные
связи, соединяющие А с Т и G с С, неодинаково прочны. Связи эти имеют
в основном электростатическую природу. В их образовании участвуют ОН- и NH2-группы.
О и N-сильно электроотрицательные элементы; они оттягивают электроны и сообщают
связанному с ними водороду положительный заряд. Положительно заряженный атом
водорода может притягиваться другими электроотрицательными группами с
неподеленными парами электронов, и в этом случае образуется водородная связь.
Прочность ее зависит от кислотности атома водорода и основности
атома-акцептора. Водородные связи прочнее межмолекулярных сил Ван-дер-Ваальса:
энергия связи может достигать 38 кДж (9 ккал) на 1 моль; в среднем это лишь не
намного больше, чем энергия тепловых колебаний при 37°С. Как видно из рис.
2.11, между гуанином и цитозином имеются три, а между тимином и аденином - две
водородные связи. Из-за малой энергии связи такие воздействия, как повышение
температуры, незначительные изменения концентрации магния или добавление
мочевины, могут приводить к большим изменениям и даже разрыву связи. При
повышении темпера туры происходит разрыв водородных связей и расхождение
полинуклеотидных цепей. Такое разрушение вторичной структуры ДНК сопровождается
поглощением света (при длине волны 259 нм) (гиперхромным эффектом).
Температуру, при которой прирост экстинкции достигает половины максимальной
величины, называют точкой плавления (Тп). Точка плавления тем выше,
чем больше в ДНК гуанина и цитозина - оснований, соединенных между собой тремя
водородными связями. Поэтому точка плавления выделенной и очищенной ДНК (рис.
2.12) служит показателем, позволяющим легко определить относительное содержание
в ней цитозина и гуанина. Содержание пар GC-это отношение суммы
молей гуанина и цитозина к сумме молей всех четырех оснований в дан ной ДНК (в
процентах). 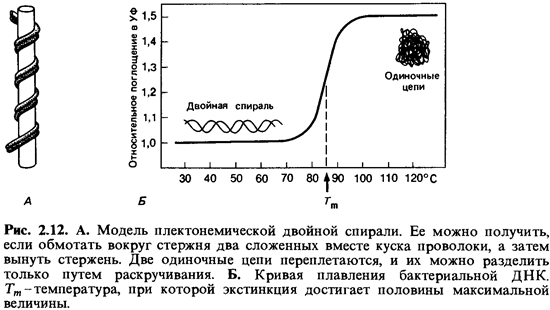
По содержанию
GC в ДНК бактерии очень сильно различаются между собой. Эта величина может
варьировать в пределах от 30% (у некоторых стафилококков и представителей
группы Cytophaga) до более чем 70% (у представителей
рода Micrococcus и некоторых миксобактерий, образующих
плодовые тела). Содержание GC видоспецифично и рассматривается как
таксономический признак.
Репликация
ДНК. ДНК содержит генетическую информацию клетки. Поэтому удвоение ДНК, всегда
предшествующее клеточному делению, должно приводить к образованию двух
совершенно идентичных хромосом. На первый взгляд этот процесс - идентичная
редупликация, или репликация, ДНК-представляется очень простым: двум цепям
достаточно только разойтись, чтобы вдоль каждой из одиночных полинуклеотидных
цепей против комплементарных оснований начали выстраиваться свободные
нуклеотиды, которым после этого останется лишь связаться между собой в цепь.
Одна из главных трудностей состоит в том, как представить себе расхождение двух
исходных цепей. Согласно данным рентгеноструктурного анализа, ДНК представляет
собой плектонемическую, а не паранемическую двойную спираль. Для того чтобы
выяснить, необходимо ли вообще раскручивание спирали, в качестве эвристических
гипотез были рассмотрены три принципиально возможных способа репликации ДНК
(рис. 2.13) (Дельбрюк и Стент, 1957): 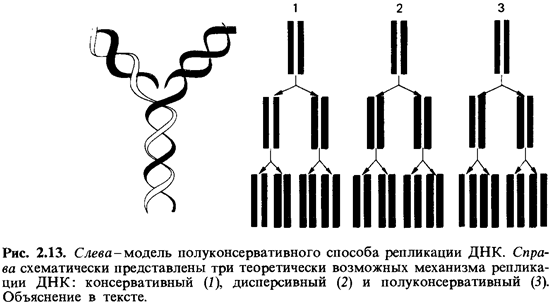
1. Консервативный
(сохраняющий) механизм. Раскручивания спирали не происходит;
родительская двойная спираль служит матрицей для синтеза двух новых цепей.
Дочерняя двойная спираль строится целиком из нового материала, а родительская
как таковая сохраняется.
2. Дисперсивный
механизм. Родительская спираль при удвоении разрывается на каждом
полуобороте путем множественной фрагментации;
3. Полуконсервативный
механизм. Родительская двойная спираль раскручивается, и на каждой
полинуклеотидной цепи образуется новая комплементарная цепь. Таким образом,
новая двойная спираль оказывается «гибридом» старой и вновь синтезированной
цепей.
С целью
выяснить, какая из этих трех возможностей соответствует действительности,
Меселсон и Сталь провели эксперименты с использованием метки и последующим
разделением меченой ДНК в градиенте плотности (рис. 2.14). 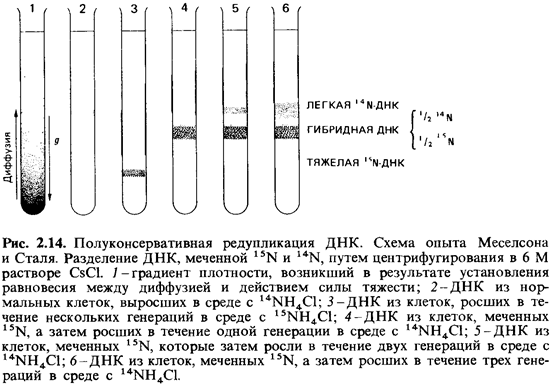
Если
центрифугировать 6 М
раствор хлористого цезия в течение многих часов при 100000 д, устанавливается
градиент плотности (равновесие между действием центробежной силы и диффузией).
Если этот раствор содержит ДНК, она скапливается в зоне, положение которой в
градиенте соответствует ее удельному весу. ДНК, выделенная из бактерий, которые
росли в присутствии 15NH4C1, оказывается на 0,8%
тяжелее обычной14N-ДНК.
Поэтому такая ДНК образует в градиенте CsCl особую полосу. Бактерию Escherichia coli на
протяжении многих поколений выращивали на 15N в качестве
источника азота, так что ее ДНК содержала только 15N. Затем к
питательной среде прибавляли в избытке 14N. До и после этого
добавления брали пробы: экстрагировали ДНК и анализировали ее в градиенте
плотности CsCl. После одного деления клеток в среде с 14N вся
ДНК имела промежуточную плотность («полутяжелая», или «гибридная», ДНК). Общее
количество такой полутяжелой ДНК оставалось постоянным на протяжении нескольких
поколений, тогда как количество легкой ДНК возрастало. Для доказательства того,
что одна цепь полутяжелой ДНК действительно содержала 15N, а
другая -14N, полутяжелую ДНК подвергали «плавлению» (нагревали до
100°С) и быстро охлаждали. После этого в градиенте плотности CsCl появлялись
две полосы, из которых одна была типична для одноцепочечной ДНК с 14N,
а другая - для такой же ДНК с 15N.
Результаты
этих опытов не совместимы ни с консервативным, ни с дисперсивным механизмом
репликации ДНК; они согласуются с представлением о полукоисервативном
механизме. При репликации цепи раскручиваются и расходятся. На каждой
одиночной цепи происходит синтез новой комплементарной цепи. В этом процессе
участвуют ДНК-полимеразы. Функция их проста: они связывают между собой
нуклеотиды, расположившиеся путем спаривания оснований в правильном порядке, и
таким образом синтезируют новую полинуклеотидную цепь. Биохимические механизмы
показаны на рис. 2.15 и поясняются в подписи к этому рисунку. С подробностями
можно ознакомиться в учебниках по молекулярной биологии.
Синтез одной
полинуклеотидной цепи таким способом объяснить не трудно: ДНК-полимераза
способна непрерывно связывать нуклеотиды по направлению от 5' к 3'-концу. Но
так как двойная спираль ДНК антипараллельна, синтез второй, противолежащей цепи
должен происходить в обратном направлении. Эти соображения и экспериментальные
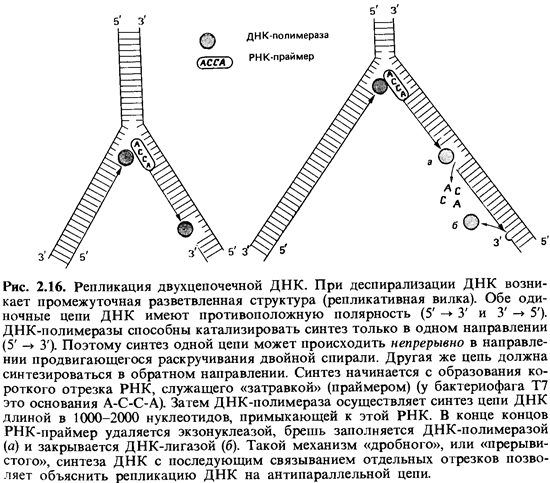
данные привели к представлению, которое иллюстрируют схемы на рис. 2.16.
Вероятно, сначала образуются только короткие (длиной около 1000 нуклеотидов)
отрезки – так называемые фрагменты Оказаки. Их синтез начинается с образования
короткой цепи РНК, которая служит «затравкой» (праймером). Затем с помощью ДНК-полимеразы
синтезируется цепь ДНК, примыкающая к РНК, длиной в 1000-2000 нуклеотидов. В
конце концов РНК-праймер удаляется, разрыв заполняется при участии другой
полимеразы, и все отрезки соединяются при помощи лигазы. Такой механизм
посегментного синтеза полинуклеотидов позволяет объяснить репликацию второй
цепи ДНК. Возможно даже, что таким способом реплицируются обе цепи. 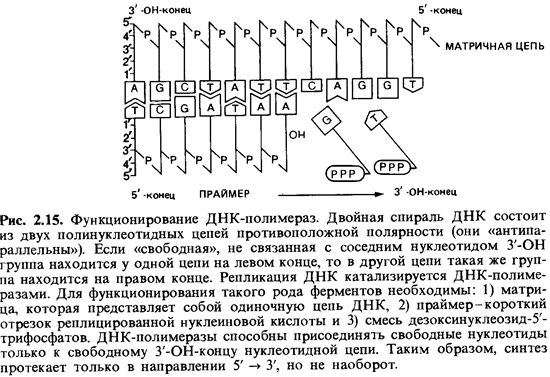
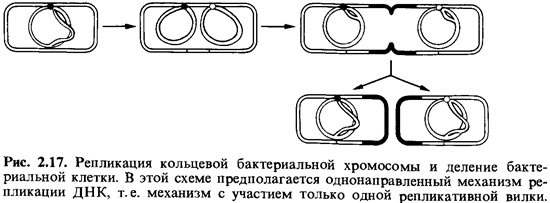
На рис. 2.17
показано, как можно представлять себе репликацию бактериальной хромосомы и
деление бактериальной клетки; в данной схеме предполагается, что репликация
всего кольца ДНК происходит в одном направлении.
Оказалось,
однако, что синтез обеих новых цепей может идти не только в одном направлении,
но и сразу в обе стороны от точки инициации. Такой механизм предполагает
раскручивание двойной спирали сразу в двух местах - с образованием двух
разветвлений репликационных вилок на одной молекуле ДНК. Удвоение хромосомы
у Е. coli занимает примерно 40 мин. Между тем эта
бактерия при благоприятных условиях делится со временем удвоения порядка всего
лишь 20 мин. Этот факт можно объяснить тем, что обе дочерние хромосомы, по
имеющимся данным, начинают новый цикл деления еще до того, как заканчивается
предыдущий. О механизме репликаций ДНК получено гораздо больше детальных
сведений, чем можно было здесь изложить. Между механизмами репликации фагов,
плазмид и бактериальных хромосом существуют значительные различия. Для
углубленного изучения этих вопросов следует обратиться к литературе по
молекулярной биологии.
Размеры
генома и число геномов. Величина генома у бактерий варьирует от вида к
виду в пределах от 0,8 до 8*106 пар оснований. Число геномов в
клетке тоже может быть у разных видов различным и зависит к тому же от условий
культивирования. В периодической культуре растущие клетки Escherichia coli имеют
от 2 до 4, Azotobacter chroococсит - от 20 до 25 и Desulfovibrio gigas
- от 10 до 15 геномов на одну клетку. (Для сравнения приведем размеры
геномов эукариот: у Neurospora crassa 19*106,
у Aspergillus niger 40*106, у человека 2,9*109 и
у Zea mays 7*109 пар оснований.)
| 



